

Mapping binding sites for the PDE4D5 cAMP-specific phosphodiesterase to the N- and C-domains of beta-arrestin using spot-immobilized peptide arrays
- PMID: 17288540
- PMCID: PMC1868836
- DOI: 10.1042/BJ20070005
Mapping binding sites for the PDE4D5 cAMP-specific phosphodiesterase to the N- and C-domains of beta-arrestin using spot-immobilized peptide arrays
Abstract
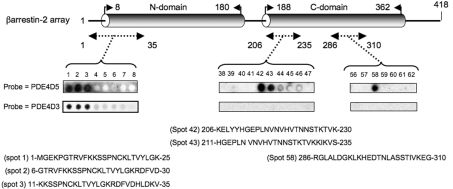
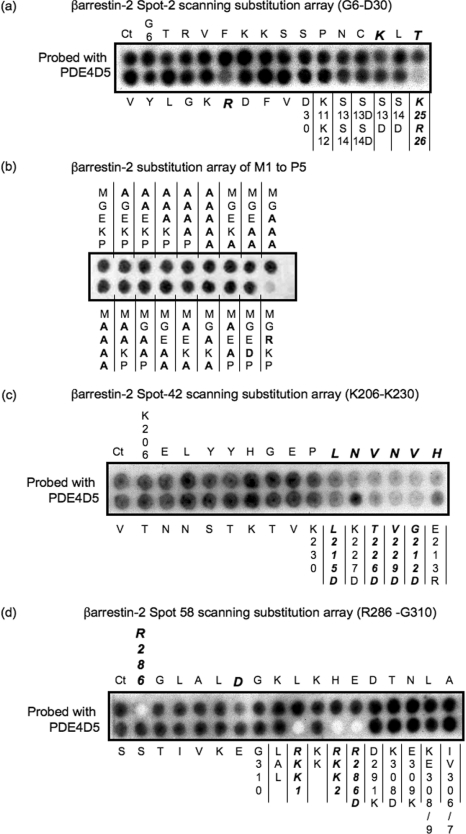
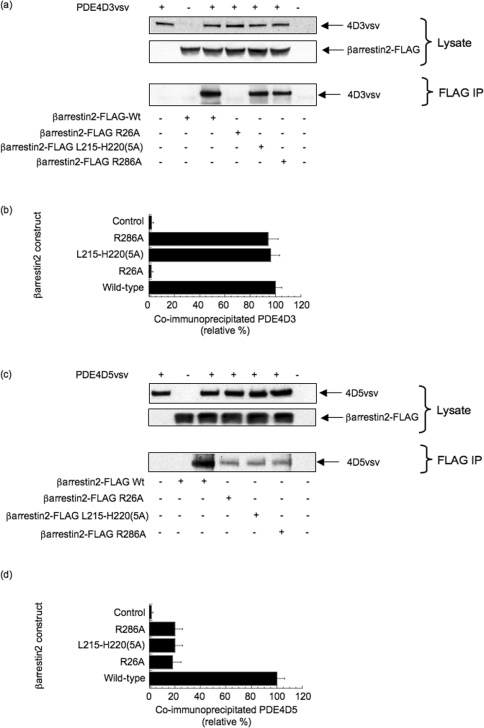
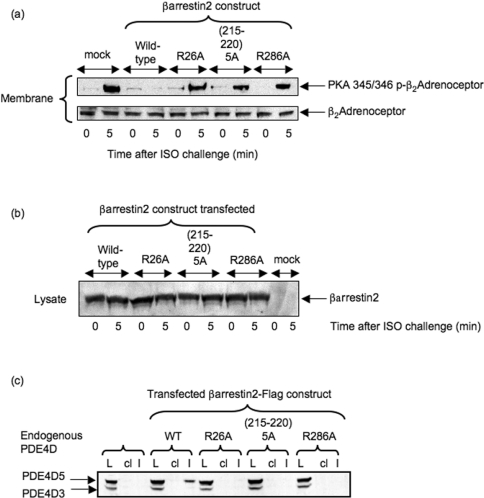
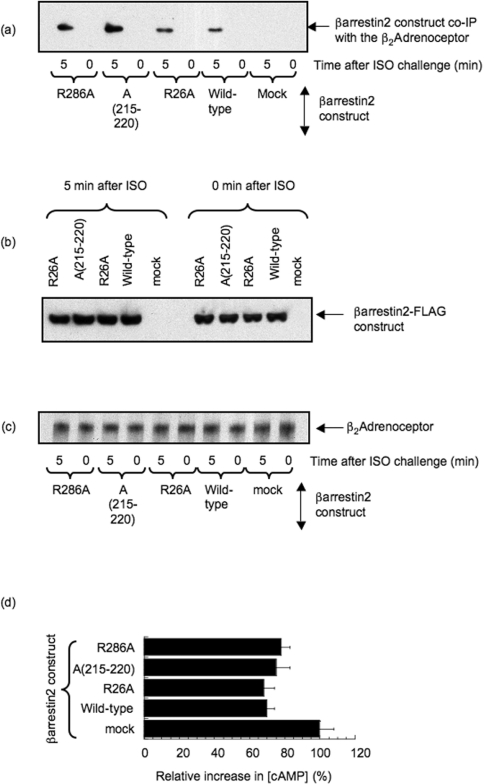
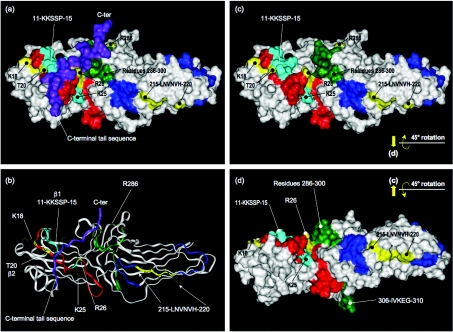
Beta2-ARs (beta2-adrenoceptors) become desensitized rapidly upon recruitment of cytosolic beta-arrestin. PDE4D5 (family 4 cAMP-specific phosphodiesterase, subfamily D, isoform 5) can be recruited in complex with beta-arrestin, whereupon it regulates PKA (cAMP-dependent protein kinase) phosphorylation of the beta2-AR. In the present study, we have used novel technology, employing a library of overlapping peptides (25-mers) immobilized on cellulose membranes that scan the entire sequence of beta-arrestin 2, to define the interaction sites on beta-arrestin 2 for binding of PDE4D5 and the cognate long isoform, PDE4D3. We have identified a binding site in the beta-arrestin 2 N-domain for the common PDE4D catalytic unit and two regions in the beta-arrestin 2 C-domain that confer specificity for PDE4D5 binding. Alanine-scanning peptide array analysis of the N-domain binding region identified severely reduced interaction with PDE4D5 upon R26A substitution, and reduced interaction upon either K18A or T20A substitution. Similar analysis of the beta-arrestin 2 C-domain identified Arg286 and Asp291, together with the Leu215-His220 region, as being important for binding PDE4D5, but not PDE4D3. Transfection with wild-type beta-arrestin 2 profoundly decreased isoprenaline-stimulated PKA phosphorylation of the beta2-AR in MEFs (mouse embryo fibroblasts) lacking both beta-arrestin 1 and beta-arrestin 2. This effect was negated using either the R26A or the R286A mutant form of beta-arrestin 2 or a mutant with substitution of an alanine cassette for Leu215-His220, which showed little or no PDE4D5 binding, but was still recruited to the beta2-AR upon isoprenaline challenge. These data show that the interaction of PDE4D5 with both the N- and C-domains of beta-arrestin 2 are essential for beta2-AR regulation.
Figures
References
-
- Tang C. M., Insel P. A. GPCR expression in the heart: “new” receptors in myocytes and fibroblasts. Trends Cardiovasc. Med. 2004;14:94–99. - PubMed
-
- Torrecilla I., Tobin A. B. Co-ordinated covalent modification of G-protein coupled receptors. Curr. Pharm. Des. 2006;12:1797–1808. - PubMed
-
- Lefkowitz R. J., Shenoy S. K. Transduction of receptor signals by β-arrestins. Science. 2005;308:512–517. - PubMed
-
- Penela P., Ribas C., Mayor F., Jr Mechanisms of regulation of the expression and function of G protein-coupled receptor kinases. Cell. Signalling. 2003;15:973–981. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous