

Checkpoint-apoptosis uncoupling in human and mouse embryonic stem cells: a source of karyotpic instability
- PMID: 17289813
- PMCID: PMC1885509
- DOI: 10.1182/blood-2006-10-054247
Checkpoint-apoptosis uncoupling in human and mouse embryonic stem cells: a source of karyotpic instability
Abstract
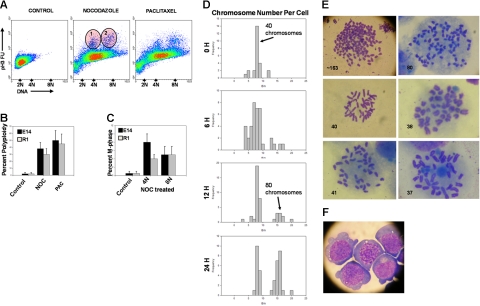
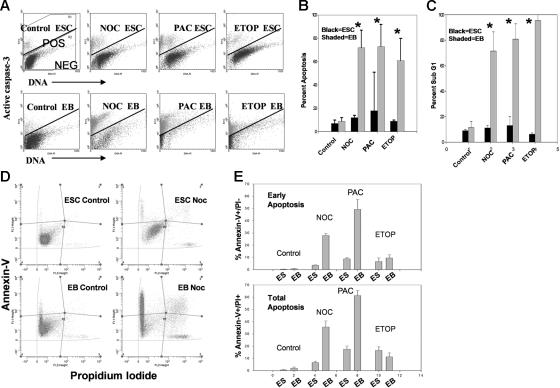
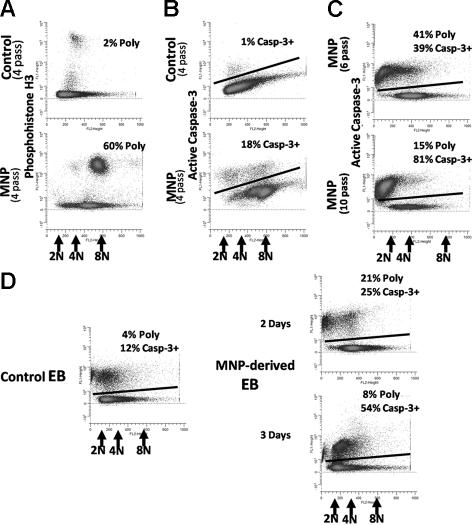
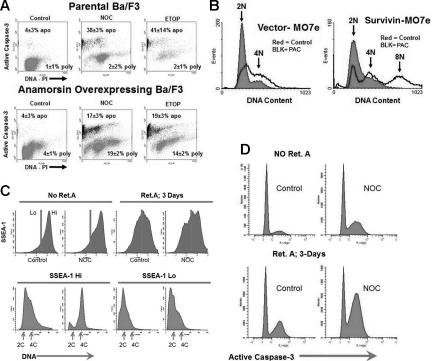
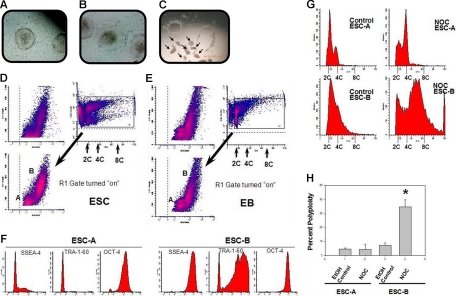
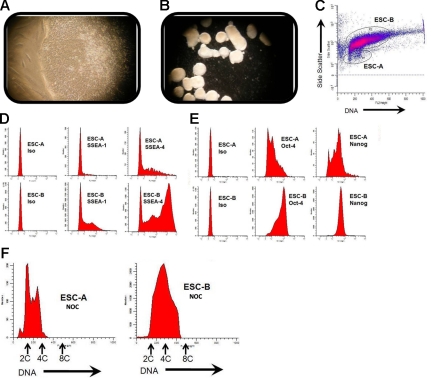
Karyotypic abnormalities in cultured embryonic stem cells (ESCs), especially near-diploid aneuploidy, are potential obstacles to ESC use in regenerative medicine. Events causing chromosomal abnormalities in ESCs may be related to events in tumor cells causing chromosomal instability (CIN) in human disease. However, the underlying mechanisms are unknown. Using multiparametric permeabilized-cell flow cytometric analysis, we found that the mitotic-spindle checkpoint, which helps maintain chromosomal integrity during all cell divisions, functions in human and mouse ESCs, but does not initiate apoptosis as it does in somatic cells. This allows an unusual tolerance to polyploidy resulting from failed mitosis, which is common in rapidly proliferating cell populations and which is reduced to near-diploid aneuploidy, which is also common in human neoplastic disease. Checkpoint activation in ESC-derived early-differentiated cells results in robust apoptosis without polyploidy/aneuploidy similar to that in somatic cells. Thus, the spindle checkpoint is "uncoupled" from apoptosis in ESCs and is a likely source of karyotypic abnormalities. This natural behavior of ESCs to tolerate/survive varying degrees of ploidy change could complicate genome-reprogramming studies and stem-cell plasticity studies, but could also reveal clues about the mechanisms of CIN in human tumors.
Figures
References
-
- Huppertz B, Herrler A. Regulation of proliferation and apoptosis during development of the preimplantation embryo and the placenta. Birth Defects Res C Embryo Today. 2005;75:249–261. - PubMed
-
- Jefford CE, Irminger-Finger I. Mechanisms of chromosome instability in cancers. Crit Rev Oncol Hematol. 2006;59:1–14. - PubMed
-
- Burdon T, Smith A, Savatier P. Signalling, cell cycle and pluripotency in embryonic stem cells. Trends Cell Biol. 2002;12:432–438. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
