

Molecular determinants for the interaction between AMPA receptors and the clathrin adaptor complex AP-2
- PMID: 17289840
- PMCID: PMC1815294
- DOI: 10.1073/pnas.0611170104
Molecular determinants for the interaction between AMPA receptors and the clathrin adaptor complex AP-2
Abstract
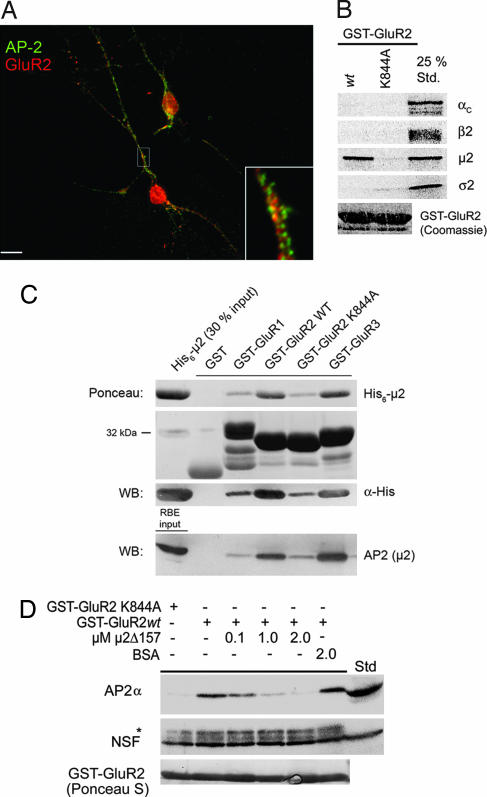
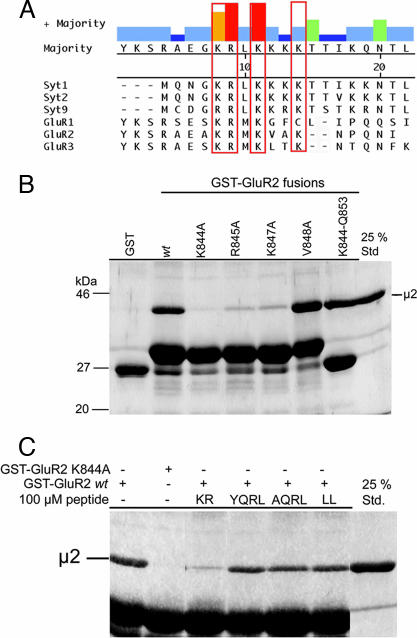
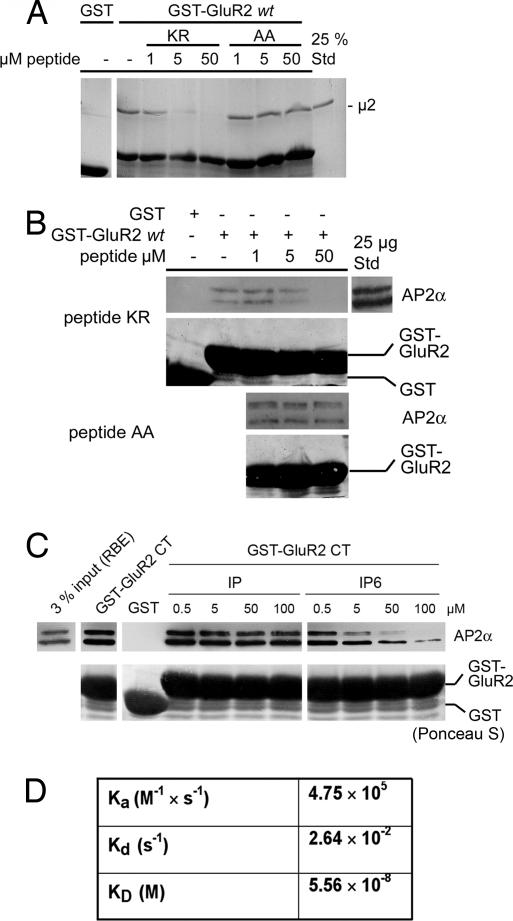
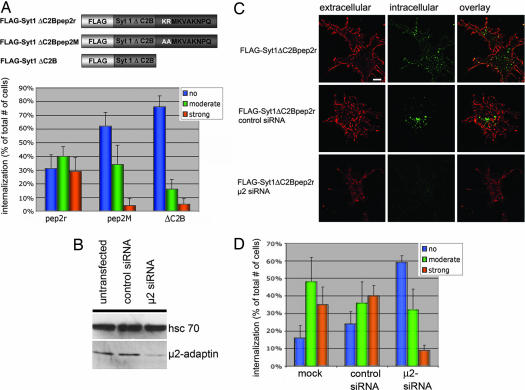
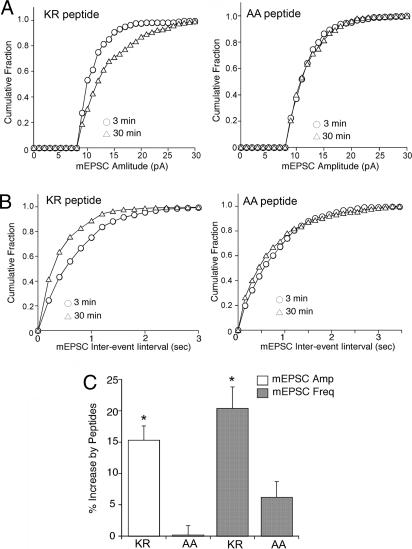
alpha-Amino-3-hydroxy-5-methylisoxazole-4-propionic acid (AMPA)-type glutamate receptors undergo constitutive and ligand-induced internalization that requires dynamin and the clathrin adaptor complex AP-2. We report here that an atypical basic motif within the cytoplasmic tails of AMPA-type glutamate receptors directly associates with mu2-adaptin by a mechanism similar to the recognition of the presynaptic vesicle protein synaptotagmin 1 by AP-2. A synaptotagmin 1-derived AP-2 binding peptide competes the interaction of the AMPA receptor subunit GluR2 with AP-2mu and increases the number of surface active glutamate receptors in living neurons. Moreover, fusion of the GluR2-derived tail peptide with a synaptotagmin 1 truncation mutant restores clathrin/AP-2-dependent internalization of the chimeric reporter protein. These data suggest that common mechanisms regulate AP-2-dependent internalization of pre- and postsynaptic membrane proteins.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
