

doi: 10.1101/gad.1521607.
ISG15 modification of the eIF4E cognate 4EHP enhances cap structure-binding activity of 4EHP
Affiliations
- PMID: 17289916
- PMCID: PMC1785121
- DOI: 10.1101/gad.1521607
Item in Clipboard
ISG15 modification of the eIF4E cognate 4EHP enhances cap structure-binding activity of 4EHP
Genes Dev.
.
Abstract
The expression of the ubiquitin-like molecule ISG15 and protein modification by ISG15 (ISGylation) are strongly activated by interferon, genotoxic stress, and pathogen infection, suggesting that ISG15 plays an important role in innate immune responses. 4EHP is an mRNA 5' cap structure-binding protein and acts as a translation suppressor by competing with eIF4E for binding to the cap structure. Here, we report that 4EHP is modified by ISG15 and ISGylated 4EHP has a much higher cap structure-binding activity. These data suggest that ISGylation of 4EHP may play an important role in cap structure-dependent translation control in immune responses.
Figures
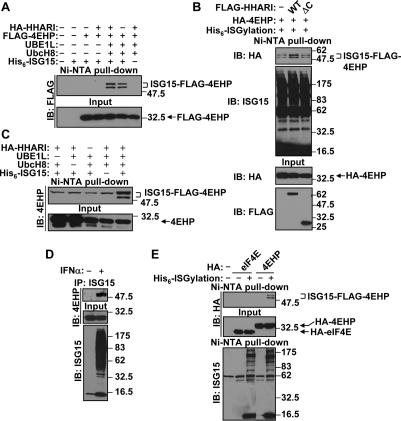
4EHP is an ISGylation target. (A) ISGylation of 4EHP. 293T cells were transfected with HA-HHARI, Flag-4EHP, UBE1L, UbcH8, and His6-ISG15 expression plasmids as indicated. Thirty-six hours after transfection, cell lysates were subjected to Ni-NTA pull-down and immunoblot (IB) analysis with anti-Flag antibody. The positions of unmodified and ISGylated 4EHP are shown on the right side as well as the molecular weight markers (in kilodaltons). (B) HHARI enhances ISGylation of 4EHP. Either Flag-HHARI (WT) or C-terminal-deleted Flag-HHARI (ΔC) was transfected to 293T cells with HA-4EHP and the His6-ISGylation system (UBE1L, UbcH8, and His6-ISG15). Cell lysates were subjected to Ni-NTA pull-down and immunoblot (IB) analysis. (C) ISGylation of endogenous 4EHP. 293T cells were transfected with UBE1L, UbcH8, HA-HHARI, and His6-ISG15 expression plasmids as indicated. Cell lysates were subjected to Ni-NTA pull-down and immunoblot (IB) analysis. (D) ISGylation of endogenous 4EHP dependent on IFNα treatment. KT-1dUBP43 cells were stimulated by 1000 U/mL of IFNα-2a for 48 h. The cell lysates were subjected to immunoprecipitation (IP) with anti-ISG15 antibody and immunoblot (IB) analysis. (E) eIF4E is not ISGylated. 293T cells were transfected with HA-eIF4E or HA-4EHP with or without the His6-ISGylation system as indicated. Cell lysates were subjected to Ni-NTA pull-down and immunoblot (IB) analysis. The positions of unmodified eIF4E and 4EHP are shown as well as ISGylated 4EHP on the right side.
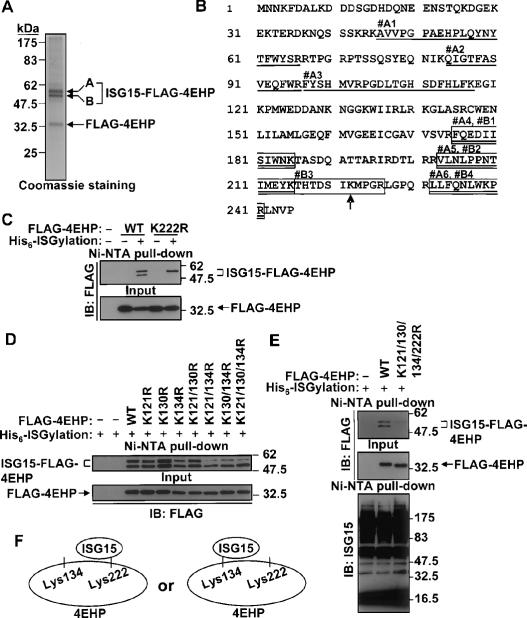
The ISGylation sites of 4EHP. (A) Coomassie staining of purified Flag-4EHP and ISGylated Flag-4EHP. Both bands A and B, which are ISGylated Flag-4EHP, were subjected to nano LC-MS/MS analysis. (B) Identified peptides by nano LC-MS/MS analysis. Peptides that came from band A are underlined (#A1–A6), and those that came from band B are boxed (#B1–B4). The lysine residue that was identified to be modified by Gly–Gly is indicated by the arrow. (C) K222R mutation eliminates the ISGylated bottom band of 4EHP. Either wild-type or K222R mutant Flag-4EHP was transfected to 293T cells with or without the His6-ISGylation system, followed by Ni-NTA pull-down and immunoblotting (IB) analysis. (D) ISGylation of Lys 134 contributes to the ISGylated top band of 4EHP. Flag-4EHP (wild type or mutants) were transfected to 293T cells with the His6-ISGylation system as indicated, followed by Ni-NTA pull-down and immunoblotting (IB) analysis. (E) K121/130/134/222R mutant 4EHP is not ISGylated. Flag-4EHP (wild type or K121/130/134/222R) was transfected to 293T cells with the His6-ISGylation system as indicated, followed by Ni-NTA pull-down and immunoblotting (IB) analysis. (F) Schematic model of the ISGylation of 4EHP. Lys 134 or Lys 222 of 4EHP can be ISGylated. The ISGylation of one of the lysine residues blocks modification on another lysine residue by structural obstruction.
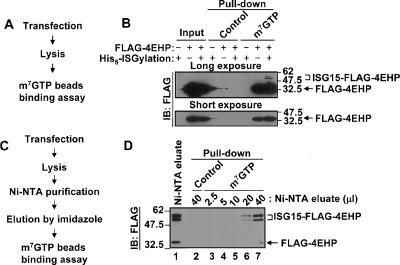
ISGylation of 4EHP enhances m7GTP cap structure-binding activity. (A) The flow chart of the direct cap structure-binding assay. (B) Flag-4EHP was transfected to 293T cells with or without His6-ISGylation system. Cell lysates were incubated with Protein A Sepharose or m7GTP-Sepharose, followed by immunoblotting (IB) analysis. Short and long exposures are shown. (C) The flow chart of the purification of ISGylated 4EHP and nonmodified 4EHP for the cap structure-binding assay. 293T cells were transfected with Flag-4EHP, HA-HHARI, and His6-ISGylation system. Cell lysates were subjected to Ni-NTA purification. The Ni-NTA-bound proteins were eluted by imidazole, and then used for the cap structure-binding assay using m7GTP beads. (D) The cap structure-binding assay. After Ni-NTA purification, the eluate fraction was subjected to immunoblot (IB) analysis to show the relative amount of Flag-4EHP and ISGylated Flag-4EHP (left, lane 1). Different amounts of Ni-NTA eluate fraction were subjected to pull-down assay with control beads or m7GTP beads as indicated (lanes 2–7). The positions of unmodified and ISGylated 4EHP are shown on the right side.
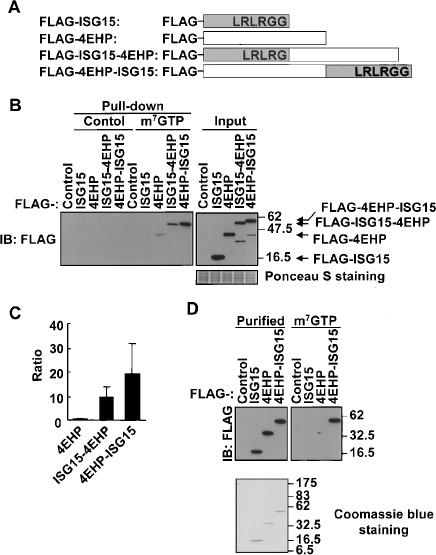
ISG15–4EHP fusion proteins have much higher m7GTP cap structure-binding activity. (A) Schematic representation of wild-type and ISG15–4EHP fusion proteins. (B) The cap structure-binding assay using wild-type or fusion proteins. 293T cells were transfected with Flag-tagged proteins as indicated, and then equal amounts of cell lysates were subjected to the cap structure-binding assay and immunoblot (IB) analysis. Ponceau S staining shows the relative amount of protein in each sample. (C) The quantification of cap structure-binding activity of 4EHP and that fused to ISG15. The amount of protein bound to m7GTP beads, which was determined by scanning densitometry of the immunoblot, was divided by the signal intensity of the input lane and normalized by that of Flag-4EHP. Data are expressed as means + standard deviations from three independent experiments. (D) Cap structure-binding assay with purified proteins. Each Flag-protein was purified from 293T cells and used for cap structure-binding assay. Coomassie staining shows no interacting protein with each purified protein.
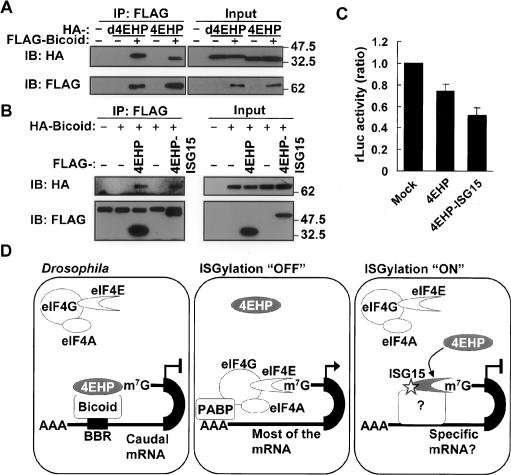
ISGylation of 4EHP enhances the mRNA-specific translation inhibition activity of 4EHP. (A) Human 4EHP binds to Drosophila Bicoid. HA-Drosophila 4EHP (d4EHP) or human 4EHP (4EHP) was transfected to 293T cells with or without Flag-Drosophila Bicoid, followed by immunoprecipitation (IP) and immunoblot (IB) analysis. (B) Human 4EHP–ISG15 fusion protein binds to Drosophila Bicoid. Flag-tagged Human 4EHP or 4EHP–ISG15 fusion protein was transfected to 293T cells with or without HA-Drosophila Bicoid, followed by immunoprecipitation (IP) and immunoblot (IB) analysis. (C) In vitro translation of capped-Renilla reniformis luciferase-Bicoid-binding region reporter mRNA with purified human 4EHP or 4EHP–ISG15 fusion protein as well as mock-purified fraction. In vitro translation of the reporter mRNA was performed in the presence of in vitro transcribed and translated Drosophila Bicoid and purified human 4EHP or 4EHP–ISG15 as well as mock-purified fraction. Data are presented as mean + standard deviation from two independent experiments, both of which were duplicated (n = 4). The values were normalized by that obtained for mock-purified fractions. (D) A model of cap structure-dependent translation regulation by ISGylated 4EHP. In Drosophila, 4EHP binds to the cap structure and Bicoid. Bicoid also binds to the Bicoid-binding region (BBR) in the 3′-untranslated region (UTR) of Caudal mRNA. This protein–mRNA complex prevents the interaction between eIF4E and the cap structure. As a result, the translation of Caudal is inhibited. Under normal conditions (ISGylation “OFF”), the eIF4F complex—which contains eIF4A, eIF4E1, and eIF4G—translates mRNA in a cap structure-dependent manner in cooperation with poly(A)-binding protein (PABP). Under a particular condition (ISGylation “ON”) that induces the ISGylation system, 4EHP is ISGylated, which leads to its stronger cap structure-binding activity to specific mRNA templates in cooperation with certain currently unknown factors. As a result, a lower amount of eIF4F complex binds to the cap structure of these mRNAs to initiate translation.
References
-
- Cho P.F., Poulin F., Cho-Park Y.A., Cho-Park I.B., Chicoine J.D., Lasko P., Sonenberg N., Poulin F., Cho-Park Y.A., Cho-Park I.B., Chicoine J.D., Lasko P., Sonenberg N., Cho-Park Y.A., Cho-Park I.B., Chicoine J.D., Lasko P., Sonenberg N., Cho-Park I.B., Chicoine J.D., Lasko P., Sonenberg N., Chicoine J.D., Lasko P., Sonenberg N., Lasko P., Sonenberg N., Sonenberg N. A new paradigm for translational control: Inhibition via 5′–3′ mRNA tethering by Bicoid and the eIF4E cognate 4EHP. Cell. 2005;121:411–423. - PubMed
-
- Dao C.T., Zhang D.E., Zhang D.E. ISG15: A ubiquitin-like enigma. Front. Biosci. 2005;10:2701–2722. - PubMed
-
- Desai S.D., Haas A.L., Wood L.M., Tsai Y.C., Pestka S., Rubin E.H., Saleem A., Nur-E-Kamal A., Liu L.F., Haas A.L., Wood L.M., Tsai Y.C., Pestka S., Rubin E.H., Saleem A., Nur-E-Kamal A., Liu L.F., Wood L.M., Tsai Y.C., Pestka S., Rubin E.H., Saleem A., Nur-E-Kamal A., Liu L.F., Tsai Y.C., Pestka S., Rubin E.H., Saleem A., Nur-E-Kamal A., Liu L.F., Pestka S., Rubin E.H., Saleem A., Nur-E-Kamal A., Liu L.F., Rubin E.H., Saleem A., Nur-E-Kamal A., Liu L.F., Saleem A., Nur-E-Kamal A., Liu L.F., Nur-E-Kamal A., Liu L.F., Liu L.F. Elevated expression of ISG15 in tumor cells interferes with the ubiquitin/26S proteasome pathway. Cancer Res. 2006;66:921–928. - PubMed
-
- Dinkova T.D., Keiper B.D., Korneeva N.L., Aamodt E.J., Rhoads R.E., Keiper B.D., Korneeva N.L., Aamodt E.J., Rhoads R.E., Korneeva N.L., Aamodt E.J., Rhoads R.E., Aamodt E.J., Rhoads R.E., Rhoads R.E. Translation of a small subset of Caenorhabditis elegans mRNAs is dependent on a specific eukaryotic translation initiation factor 4E isoform. Mol. Cell. Biol. 2005;25:100–113. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous