

Redirection of sphingolipid metabolism toward de novo synthesis of ethanolamine in Leishmania
- PMID: 17290222
- PMCID: PMC1852826
- DOI: 10.1038/sj.emboj.7601565
Redirection of sphingolipid metabolism toward de novo synthesis of ethanolamine in Leishmania
Abstract
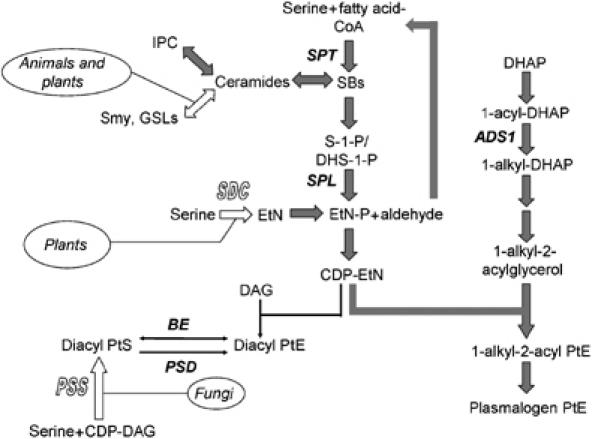
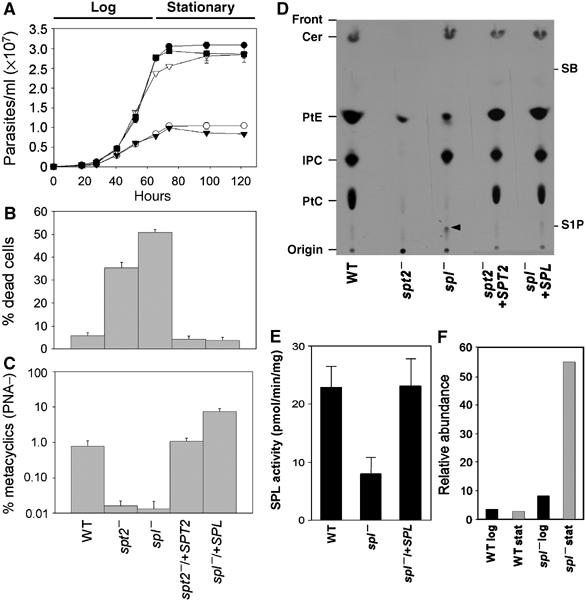
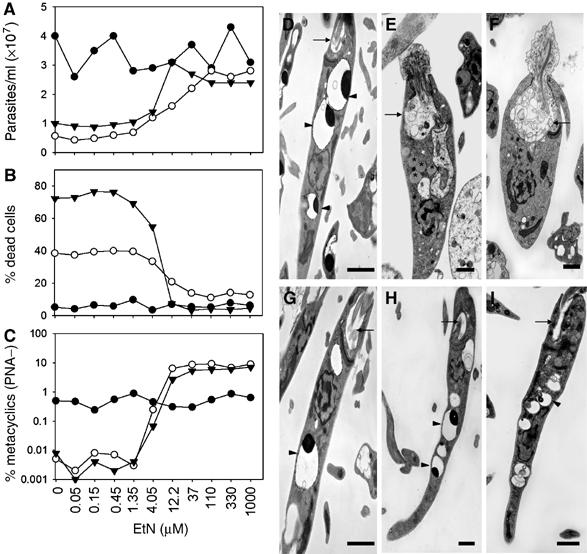
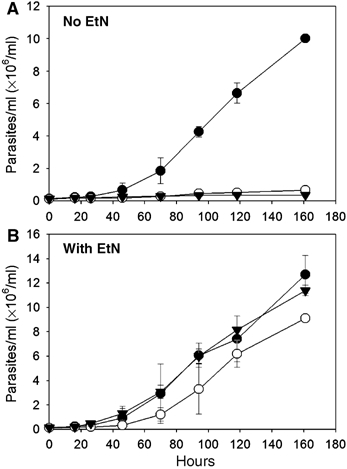
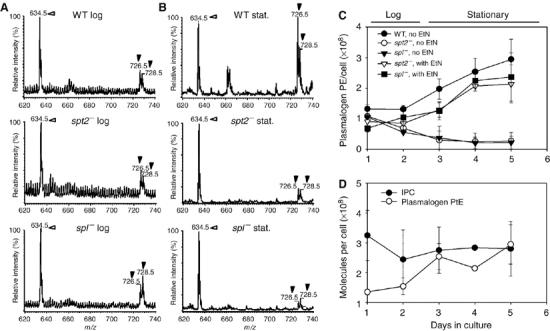
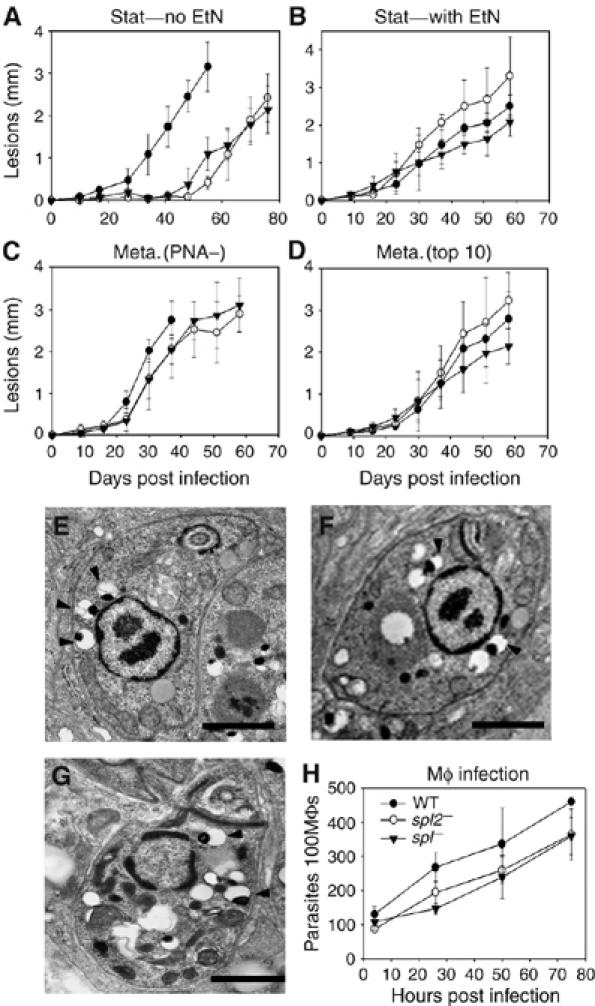
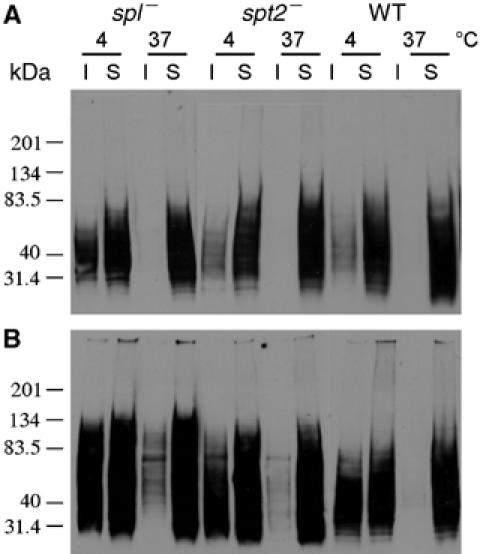
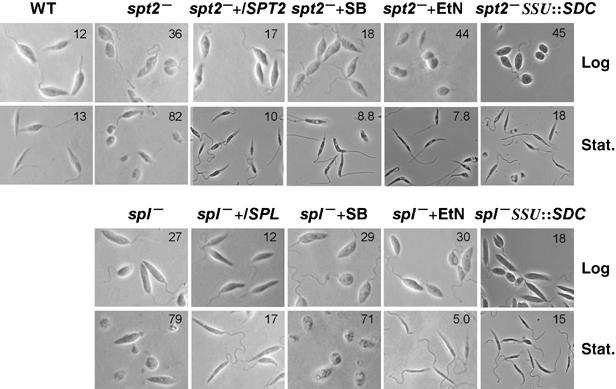
In most eukaryotes, sphingolipids (SLs) are critical membrane components and signaling molecules. However, mutants of the trypanosomatid protozoan Leishmania lacking serine palmitoyltransferase (spt2-) and SLs grow well, although they are defective in stationary phase differentiation and virulence. Similar phenotypes were observed in sphingolipid (SL) mutant lacking the degradatory enzyme sphingosine 1-phosphate lyase (spl-). This epistatic interaction suggested that a metabolite downstream of SLs was responsible. Here we show that unlike other organisms, the Leishmania SL pathway has evolved to be the major route for ethanolamine (EtN) synthesis, as EtN supplementation completely reversed the viability and differentiation defects of both mutants. Thus Leishmania has undergone two major metabolic shifts: first in de-emphasizing the metabolic roles of SLs themselves in growth, signaling, and maintenance of membrane microdomains, which may arise from the unique combination of abundant parasite lipids; Second, freed of typical SL functional constraints and a lack of alternative routes to produce EtN, Leishmania redirected SL metabolism toward bulk EtN synthesis. Our results thus reveal a striking example of remodeling of the SL metabolic pathway in Leishmania.
Figures
Similar articles
-
Sphingolipids are essential for differentiation but not growth in Leishmania.EMBO J. 2003 Nov 17;22(22):6016-26. doi: 10.1093/emboj/cdg584. EMBO J. 2003. PMID: 14609948 Free PMC article.
-
Degradation of host sphingomyelin is essential for Leishmania virulence.PLoS Pathog. 2009 Dec;5(12):e1000692. doi: 10.1371/journal.ppat.1000692. Epub 2009 Dec 11. PLoS Pathog. 2009. PMID: 20011126 Free PMC article.
-
Sphingolipid-free Leishmania are defective in membrane trafficking, differentiation and infectivity.Mol Microbiol. 2004 Apr;52(2):313-27. doi: 10.1111/j.1365-2958.2003.03975.x. Mol Microbiol. 2004. PMID: 15066023
-
Phospholipid and sphingolipid metabolism in Leishmania.Mol Biochem Parasitol. 2010 Apr;170(2):55-64. doi: 10.1016/j.molbiopara.2009.12.004. Epub 2009 Dec 23. Mol Biochem Parasitol. 2010. PMID: 20026359 Free PMC article. Review.
-
The sphingolipid anteome: implications for evolution of the sphingolipid metabolic pathway.FEBS Lett. 2022 Sep;596(18):2345-2363. doi: 10.1002/1873-3468.14457. Epub 2022 Aug 4. FEBS Lett. 2022. PMID: 35899376 Review.
Cited by
-
Ethanolaminephosphate cytidyltransferase is essential for survival, lipid homeostasis and stress tolerance in Leishmania major.bioRxiv [Preprint]. 2023 Jan 11:2023.01.10.523530. doi: 10.1101/2023.01.10.523530. bioRxiv. 2023. Update in: PLoS Pathog. 2023 Jul 28;19(7):e1011112. doi: 10.1371/journal.ppat.1011112. PMID: 36712124 Free PMC article. Updated. Preprint.
-
De novo sphingolipid synthesis is essential for viability, but not for transport of glycosylphosphatidylinositol-anchored proteins, in African trypanosomes.Eukaryot Cell. 2007 Mar;6(3):454-64. doi: 10.1128/EC.00283-06. Epub 2007 Jan 12. Eukaryot Cell. 2007. PMID: 17220466 Free PMC article.
-
Phosphatidylcholine synthesis through cholinephosphate cytidylyltransferase is dispensable in Leishmania major.Sci Rep. 2019 May 20;9(1):7602. doi: 10.1038/s41598-019-44086-6. Sci Rep. 2019. PMID: 31110206 Free PMC article.
-
Characterization of inositol phosphorylceramides from Leishmania major by tandem mass spectrometry with electrospray ionization.J Am Soc Mass Spectrom. 2007 Sep;18(9):1591-604. doi: 10.1016/j.jasms.2007.05.017. Epub 2007 Jun 2. J Am Soc Mass Spectrom. 2007. PMID: 17627842 Free PMC article.
-
Lipid metabolism in Trypanosoma brucei.Mol Biochem Parasitol. 2010 Aug;172(2):66-79. doi: 10.1016/j.molbiopara.2010.04.001. Epub 2010 Apr 9. Mol Biochem Parasitol. 2010. PMID: 20382188 Free PMC article. Review.
References
-
- Araujo-Santos JM, Gamarro F, Castanys S, Herrmann A, Pomorski T (2003) Rapid transport of phospholipids across the plasma membrane of Leishmania infantum. Biochem Biophys Res Commun 306: 250–255 - PubMed
-
- Berriman M, Ghedin E, Hertz-Fowler C, Blandin G, Renauld H, Bartholomeu DC, Lennard NJ, Caler E, Hamlin NE, Haas B, Bohme U, Hannick L, Aslett MA, Shallom J, Marcello L, Hou L, Wickstead B, Alsmark UC, Arrowsmith C, Atkin RJ, Barron AJ, Bringaud F, Brooks K, Carrington M, Cherevach I, Chillingworth TJ, Churcher C, Clark LN, Corton CH, Cronin A, Davies RM, Doggett J, Djikeng A, Feldblyum T, Field MC, Fraser A, Goodhead I, Hance Z, Harper D, Harris BR, Hauser H, Hostetler J, Ivens A, Jagels K, Johnson D, Johnson J, Jones K, Kerhornou AX, Koo H, Larke N, Landfear S, Larkin C, Leech V, Line A, Lord A, Macleod A, Mooney PJ, Moule S, Martin DM, Morgan GW, Mungall K, Norbertczak H, Ormond D, Pai G, Peacock CS, Peterson J, Quail MA, Rabbinowitsch E, Rajandream MA, Reitter C, Salzberg SL, Sanders M, Schobel S, Sharp S, Simmonds M, Simpson AJ, Tallon L, Turner CM, Tait A, Tivey AR, Van Aken S, Walker D, Wanless D, Wang S, White B, White O, Whitehead S, Woodward J, Wortman J, Adams MD, Embley TM, Gull K, Ullu E, Barry JD, Fairlamb AH, Opperdoes F, Barrell BG, Donelson JE, Hall N, Fraser CM, Melville SE, El-Sayed NM (2005) The genome of the African trypanosome Trypanosoma brucei. Science 309: 416–422 - PubMed
-
- Besteiro S, Williams RA, Morrison LS, Coombs GH, Mottram JC (2006) Endosome sorting and autophagy are essential for differentiation and virulence of Leishmania major. J Biol Chem 281: 11384–11396 - PubMed
-
- Cruz A, Beverley SM (1990) Gene replacement in parasitic protozoa. Nature 348: 171–173 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
