

No detectable effect of RNA-binding protein Hfq absence in Staphylococcus aureus
- PMID: 17291347
- PMCID: PMC1800855
- DOI: 10.1186/1471-2180-7-10
No detectable effect of RNA-binding protein Hfq absence in Staphylococcus aureus
Abstract
Background: The RNA-binding protein Hfq is involved in stress and virulence of several pathogens, probably due to its role as mediator in small RNA (sRNA)-mRNA interactions. In this study, we investigate the function of Hfq in the Gram-positive pathogen Staphylococcus aureus, by constructing hfq null mutant derivatives.
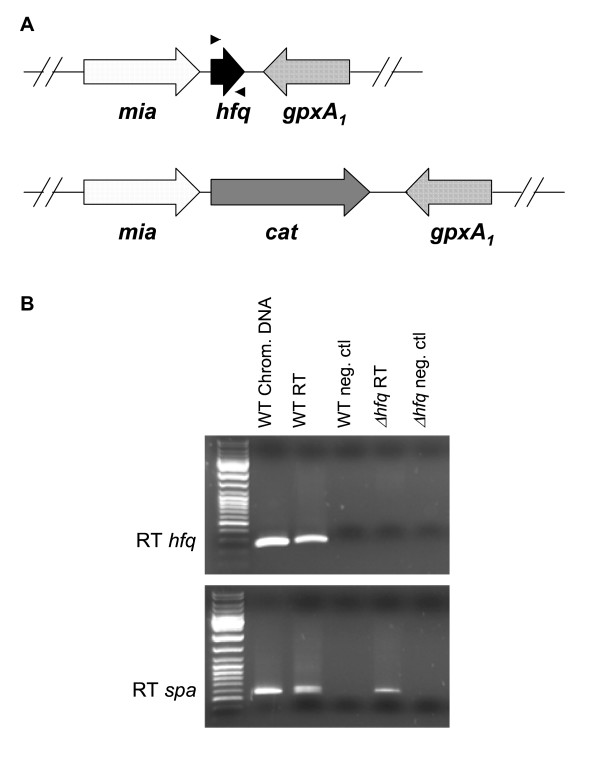
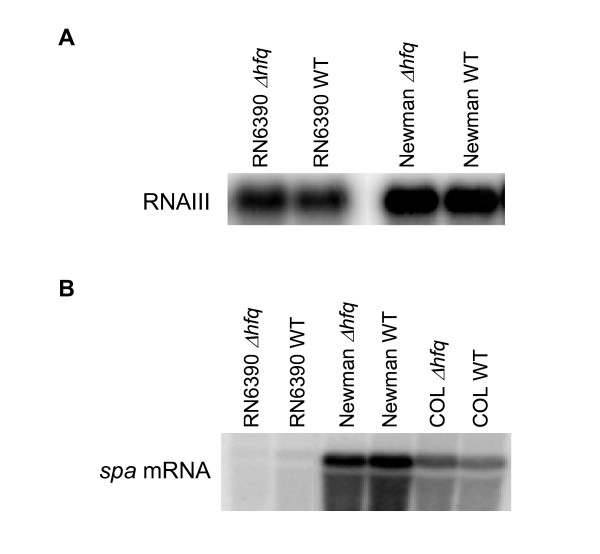
Results: We report that unexpectedly, in S. aureus, Hfq does not seem to play a crucial role in stress response, RNAIII or spa mRNA quantity and exoprotein expression, as tested in three virulent genetic backgrounds. Moreover, a global analysis of the RN6390 hfq mutant, which tests approximately 2000 phenotypes, supports our results concerning the non-implication of Hfq in stress response, and shows that Hfq is also not involved in resistance to several chemical agents and antibiotics and does not seem to be implicated in metabolic pathways.
Conclusion: Our data suggest that although sRNA-mRNA interactions in S. aureus are decisive for gene expression regulation, they do not require the RNA-chaperone protein Hfq. These interactions possibly require an RNA-chaperone protein other than Hfq, which remains to be found.
Figures

Similar articles
-
RNAIII activates map expression by forming an RNA-RNA complex in Staphylococcus aureus.FEBS Lett. 2011 Mar 23;585(6):899-905. doi: 10.1016/j.febslet.2011.02.021. Epub 2011 Feb 22. FEBS Lett. 2011. PMID: 21349272
-
Molecular modeling study on the effect of residues distant from the nucleotide-binding portion on RNA binding in Staphylococcus aureus Hfq.J Mol Graph Model. 2009 Oct;28(3):253-60. doi: 10.1016/j.jmgm.2009.08.003. Epub 2009 Aug 8. J Mol Graph Model. 2009. PMID: 19748809
-
Lack of interchangeability of Hfq-like proteins.Biochimie. 2012 Jul;94(7):1554-9. doi: 10.1016/j.biochi.2012.01.016. Epub 2012 Feb 2. Biochimie. 2012. PMID: 22326874
-
[The regulatory role of the Hfq protein in bacterial cells].Mol Biol (Mosk). 2002 Nov-Dec;36(6):970-7. Mol Biol (Mosk). 2002. PMID: 12500533 Review. Russian.
-
Contributions of the Hfq protein to translation regulation by small noncoding RNAs binding to the mRNA coding sequence.Acta Biochim Pol. 2016;63(4):701-707. doi: 10.18388/abp.2016_1362. Epub 2016 Nov 23. Acta Biochim Pol. 2016. PMID: 27878140 Review.
Cited by
-
Global small RNA chaperone Hfq and regulatory small RNAs are important virulence regulators in Erwinia amylovora.J Bacteriol. 2013 Apr;195(8):1706-17. doi: 10.1128/JB.02056-12. Epub 2013 Feb 1. J Bacteriol. 2013. PMID: 23378513 Free PMC article.
-
Staphylococcus aureus RNAIII coordinately represses the synthesis of virulence factors and the transcription regulator Rot by an antisense mechanism.Genes Dev. 2007 Jun 1;21(11):1353-66. doi: 10.1101/gad.423507. Genes Dev. 2007. PMID: 17545468 Free PMC article.
-
A multicopy sRNA of Listeria monocytogenes regulates expression of the virulence adhesin LapB.Nucleic Acids Res. 2014 Aug;42(14):9383-98. doi: 10.1093/nar/gku630. Epub 2014 Jul 17. Nucleic Acids Res. 2014. PMID: 25034691 Free PMC article.
-
dRNA-Seq Reveals Genomewide TSSs and Noncoding RNAs of Plant Beneficial Rhizobacterium Bacillus amyloliquefaciens FZB42.PLoS One. 2015 Nov 5;10(11):e0142002. doi: 10.1371/journal.pone.0142002. eCollection 2015. PLoS One. 2015. PMID: 26540162 Free PMC article.
-
Tracking the Elusive Function of Bacillus subtilis Hfq.PLoS One. 2015 Apr 27;10(4):e0124977. doi: 10.1371/journal.pone.0124977. eCollection 2015. PLoS One. 2015. PMID: 25915524 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
