

Transcription reactivation steps stimulated by oocyte maturation in C. elegans
- PMID: 17291483
- PMCID: PMC1913287
- DOI: 10.1016/j.ydbio.2006.12.039
Transcription reactivation steps stimulated by oocyte maturation in C. elegans
Abstract
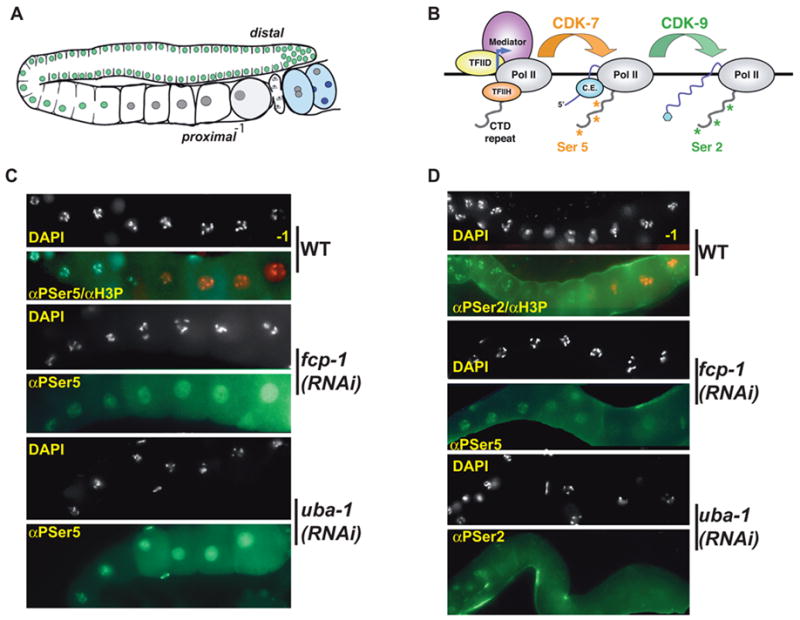
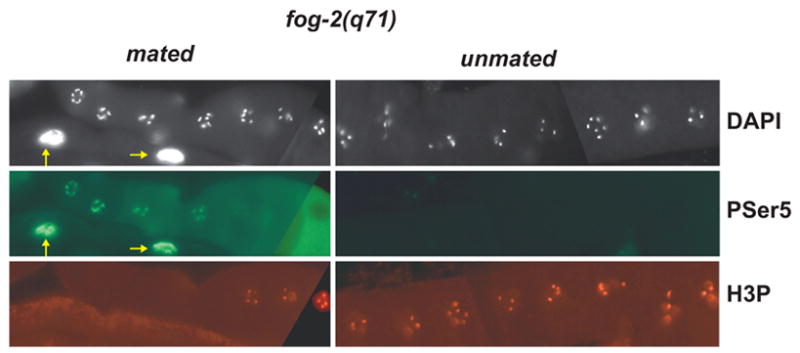
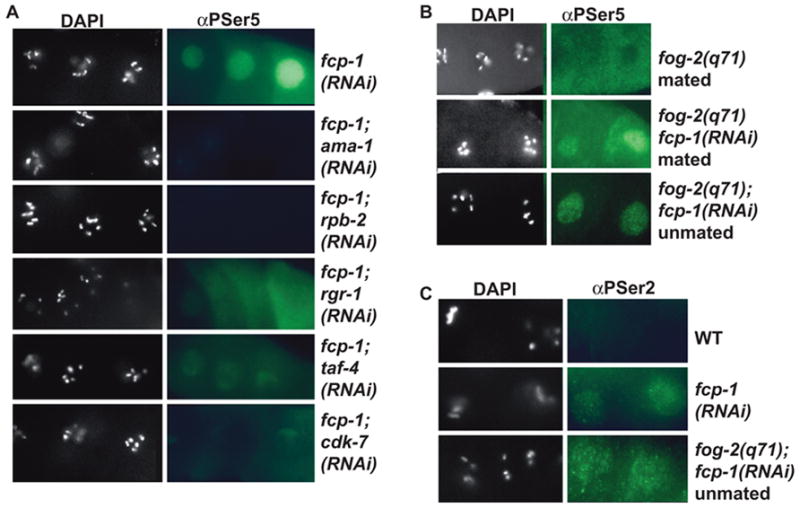
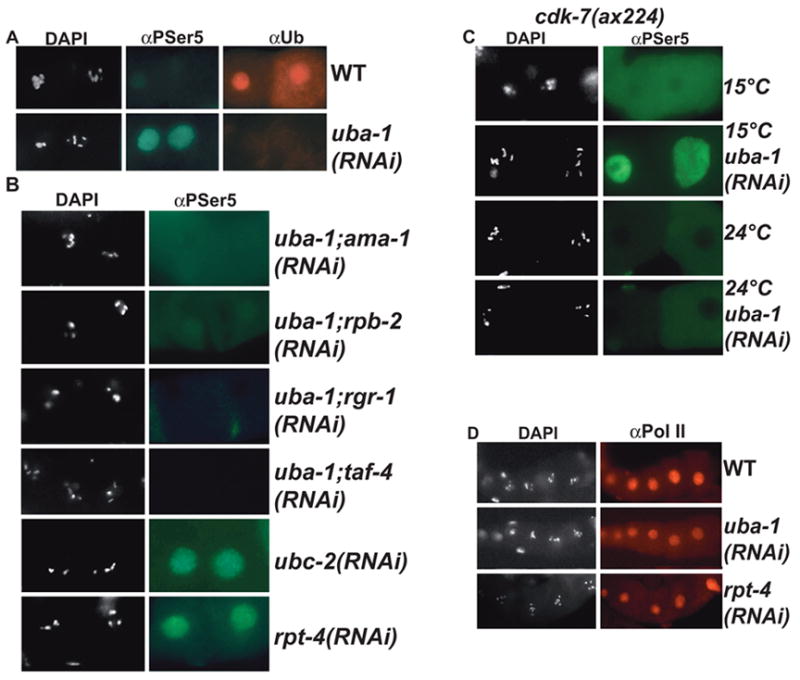
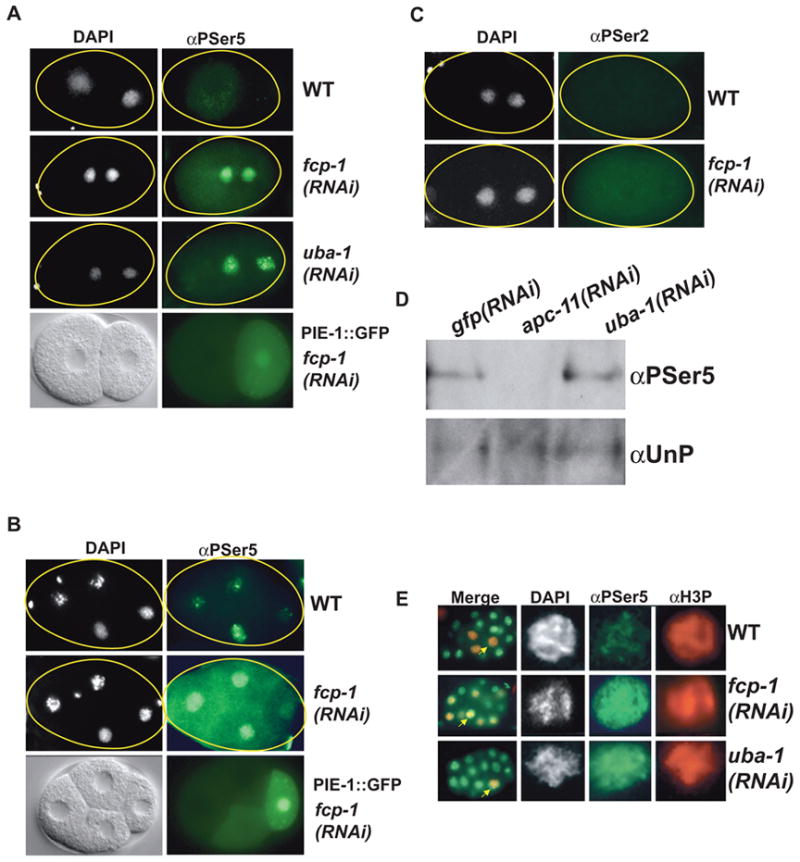
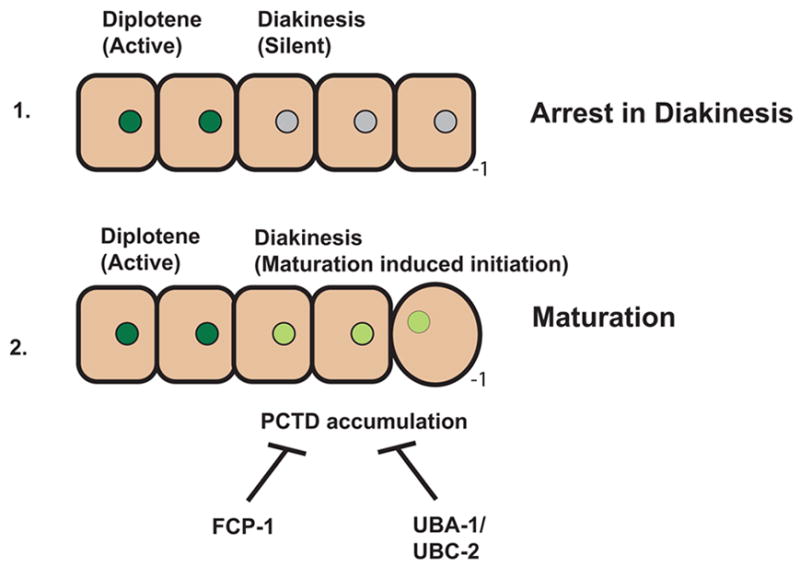
Developing oocytes produce materials that will support early embryonic development then cease transcription before fertilization. Later, a distinct transcription program is established in the embryo. Little is understood about how these global gene regulation transitions are effected. We have investigated in C. elegans how oocyte transcription is influenced by maturation, a process that releases meiotic arrest and prepares for fertilization. By monitoring transcription-associated phosphorylation of the RNA polymerase II (Pol II) C-terminal domain (CTD), we find that oocyte transcription shuts down independently of maturation. Surprisingly, maturation signals then induce CTD phosphorylation that is associated specifically with transcription initiation steps and accumulates to high levels when expression of the CTD phosphatase FCP-1 is inhibited. This CTD phosphorylation is also uncovered when a ubiquitylation pathway is blocked, or when maturation is stimulated precociously. CTD phosphorylation is similarly detected during embryonic mitosis, when transcription is also largely silenced. We conclude that oocyte maturation signals induce abortive transcription events in which FCP-1 may recycle phosphorylated Pol II and that analogous processes may occur during mitosis. Our findings suggest that maturation signals may initiate preparations for embryonic transcription, possibly as part of a broader program that begins the transition from maternal to zygotic gene expression.
Figures

References
-
- Baugh LR, Hill AA, Slonim DK, Brown EL, Hunter CP. Composition and dynamics of the Caenorhabditis elegans early embryonic transcriptome. Development. 2003;130:889–900. - PubMed
-
- Bentley DL. Rules of engagement: co-transcriptional recruitment of pre-mRNA processing factors. Curr Opin Cell Biol. 2005;17:251–6. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
