

Ribosome recycling factor and release factor 3 action promotes TnaC-peptidyl-tRNA Dropoff and relieves ribosome stalling during tryptophan induction of tna operon expression in Escherichia coli
- PMID: 17293419
- PMCID: PMC1855834
- DOI: 10.1128/JB.01868-06
Ribosome recycling factor and release factor 3 action promotes TnaC-peptidyl-tRNA Dropoff and relieves ribosome stalling during tryptophan induction of tna operon expression in Escherichia coli
Abstract
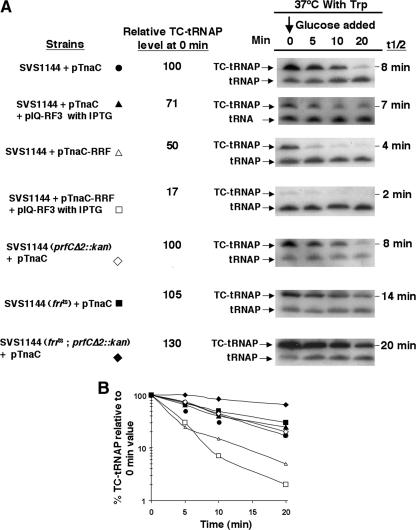
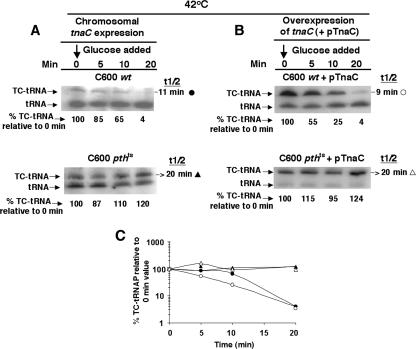
Upon tryptophan induction of tna operon expression in Escherichia coli, the leader peptidyl-tRNA, TnaC-tRNA(2)(Pro), resists cleavage, resulting in ribosome stalling at the tnaC stop codon. This stalled ribosome blocks Rho factor binding and action, preventing transcription termination in the tna operon's leader region. Plasmid-mediated overexpression of tnaC was previously shown to inhibit cell growth by reducing uncharged tRNA(2)(Pro) availability. Which factors relieve ribosome stalling, facilitate TnaC-tRNA(2)(Pro) cleavage, and relieve growth inhibition were addressed in the current study. In strains containing the chromosomal tna operon and lacking a tnaC plasmid, the overproduction of ribosome recycling factor (RRF) and release factor 3 (RF3) reduced tna operon expression. Their overproduction in vivo also increased the rate of cleavage of TnaC-tRNA(2)(Pro), relieving the growth inhibition associated with plasmid-mediated tnaC overexpression. The overproduction of elongation factor G or initiation factor 3 did not have comparable effects, and tmRNA was incapable of attacking TnaC-tRNA(2)(Pro) in stalled ribosome complexes. The stability of TnaC-tRNA(2)(Pro) was increased appreciably in strains deficient in RRF and RF3 or deficient in peptidyl-tRNA hydrolase. These findings reveal the existence of a natural mechanism whereby an amino acid, tryptophan, binds to ribosomes that have just completed the synthesis of TnaC-tRNA(2)(Pro). Bound tryptophan inhibits RF2-mediated cleavage of TnaC-tRNA(2)(Pro), resulting in the stalling of the ribosome translating tnaC mRNA. This stalling results in increased transcription of the structural genes of the tna operon. RRF and RF3 then bind to this stalled ribosome complex and slowly release TnaC-tRNA(2)(Pro). This release allows ribosome recycling and permits the cleavage of TnaC-tRNA(2)(Pro) by peptidyl-tRNA hydrolase.
Figures


Similar articles
-
Features of ribosome-peptidyl-tRNA interactions essential for tryptophan induction of tna operon expression.Mol Cell. 2005 Aug 5;19(3):333-43. doi: 10.1016/j.molcel.2005.06.013. Mol Cell. 2005. PMID: 16061180
-
Changes produced by bound tryptophan in the ribosome peptidyl transferase center in response to TnaC, a nascent leader peptide.Proc Natl Acad Sci U S A. 2006 Mar 7;103(10):3598-603. doi: 10.1073/pnas.0600082103. Epub 2006 Feb 27. Proc Natl Acad Sci U S A. 2006. PMID: 16505360 Free PMC article.
-
The mechanism of tryptophan induction of tryptophanase operon expression: tryptophan inhibits release factor-mediated cleavage of TnaC-peptidyl-tRNA(Pro).Proc Natl Acad Sci U S A. 2001 Jul 31;98(16):8997-9001. doi: 10.1073/pnas.171299298. Epub 2001 Jul 24. Proc Natl Acad Sci U S A. 2001. PMID: 11470925 Free PMC article.
-
Some novel transcription attenuation mechanisms used by bacteria.Biochimie. 1996;78(11-12):1017-24. doi: 10.1016/s0300-9084(97)86725-9. Biochimie. 1996. PMID: 9150880 Review.
-
The different roles of tryptophan transfer RNA in regulating trp operon expression in E. coli versus B. subtilis.Trends Genet. 2004 Aug;20(8):367-74. doi: 10.1016/j.tig.2004.06.007. Trends Genet. 2004. PMID: 15262409 Review.
Cited by
-
Complementary roles of initiation factor 1 and ribosome recycling factor in 70S ribosome splitting.EMBO J. 2008 Jun 18;27(12):1706-17. doi: 10.1038/emboj.2008.99. Epub 2008 May 22. EMBO J. 2008. PMID: 18497739 Free PMC article.
-
Ribosome Rescue Pathways in Bacteria.Front Microbiol. 2021 Mar 18;12:652980. doi: 10.3389/fmicb.2021.652980. eCollection 2021. Front Microbiol. 2021. PMID: 33815344 Free PMC article. Review.
-
Drop-off-reinitiation triggered by EF-G-driven mistranslocation and its alleviation by EF-P.Nucleic Acids Res. 2022 Mar 21;50(5):2736-2753. doi: 10.1093/nar/gkac068. Nucleic Acids Res. 2022. PMID: 35188576 Free PMC article.
-
Structural basis for the substrate recognition and catalysis of peptidyl-tRNA hydrolase.Nucleic Acids Res. 2012 Nov 1;40(20):10521-31. doi: 10.1093/nar/gks790. Epub 2012 Aug 25. Nucleic Acids Res. 2012. PMID: 22923517 Free PMC article.
-
Termination and antitermination: RNA polymerase runs a stop sign.Nat Rev Microbiol. 2011 May;9(5):319-29. doi: 10.1038/nrmicro2560. Epub 2011 Apr 11. Nat Rev Microbiol. 2011. PMID: 21478900 Free PMC article. Review.
References
-
- Cruz-Vera, L. R., S. Rajagopal, C. Squires, and C. Yanofsky. 2005. Features of ribosome-peptidyl-tRNA interactions essential for tryptophan induction of tna operon expression. Mol. Cell 19:333-343. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
