

Fate of cranial neural crest cells during craniofacial development in endothelin-A receptor-deficient mice
- PMID: 17294360
- PMCID: PMC2810159
- DOI: 10.1387/ijdb.062237ma
Fate of cranial neural crest cells during craniofacial development in endothelin-A receptor-deficient mice
Abstract
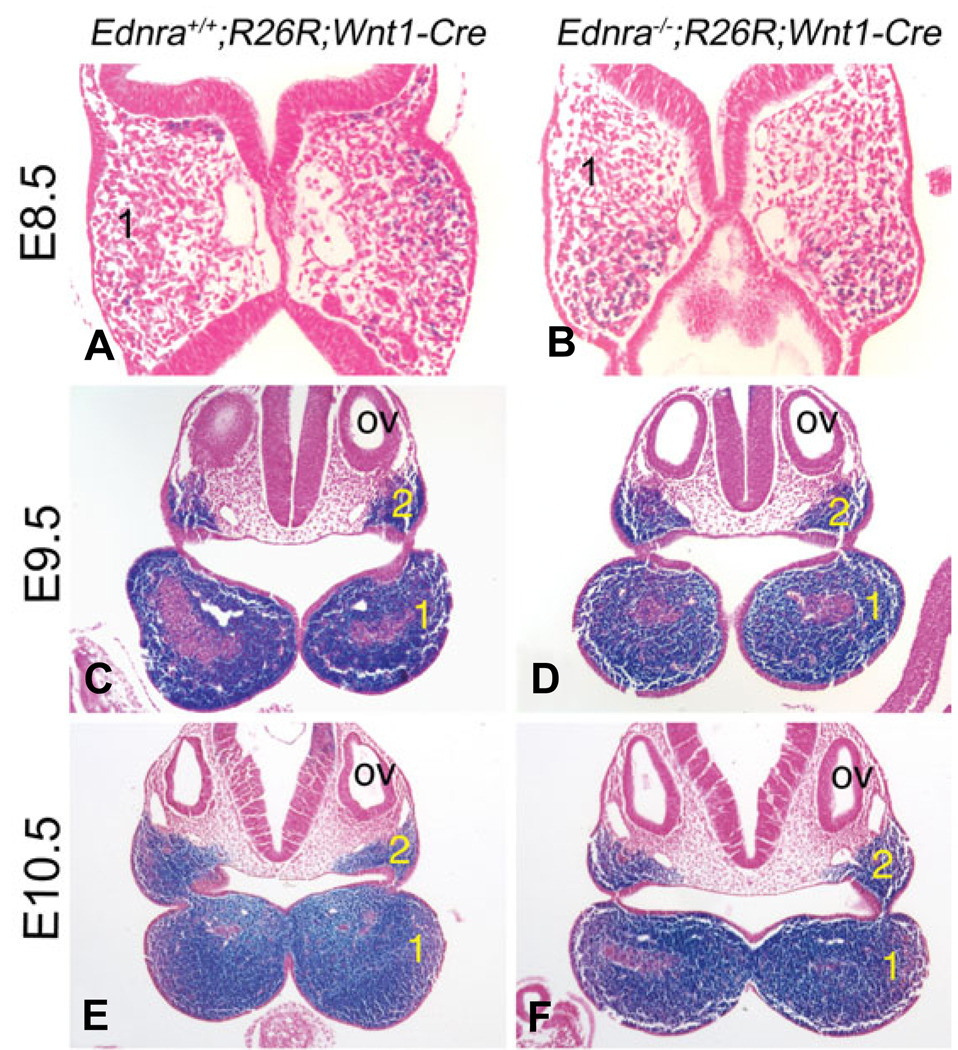
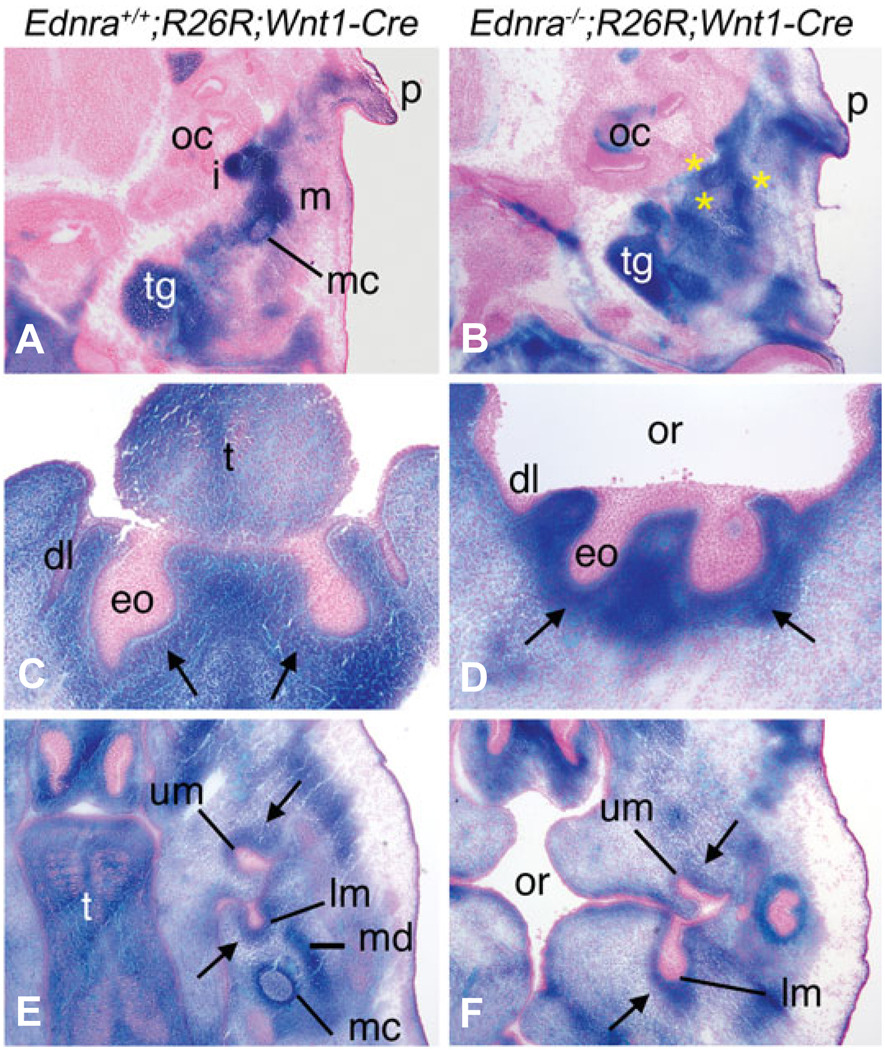
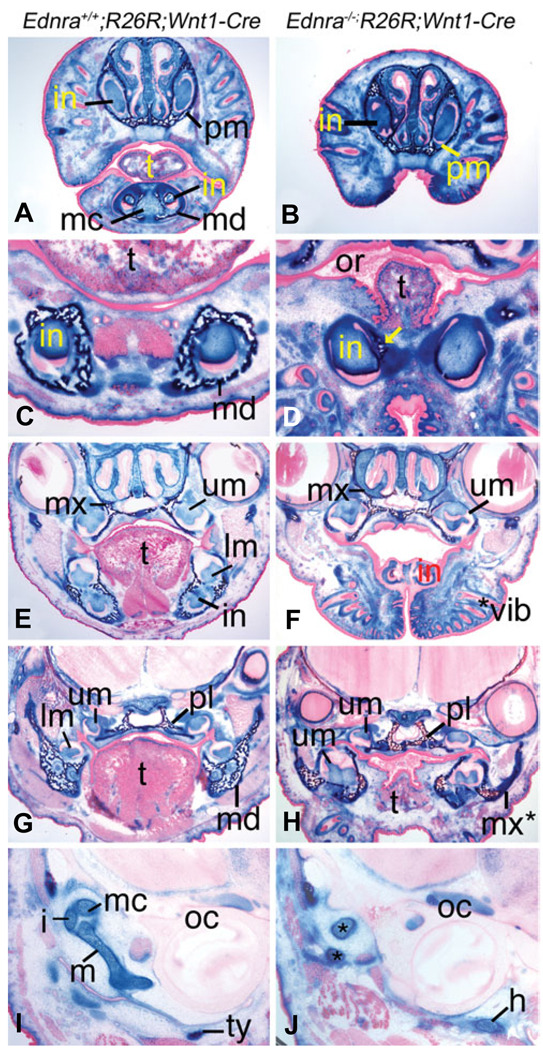
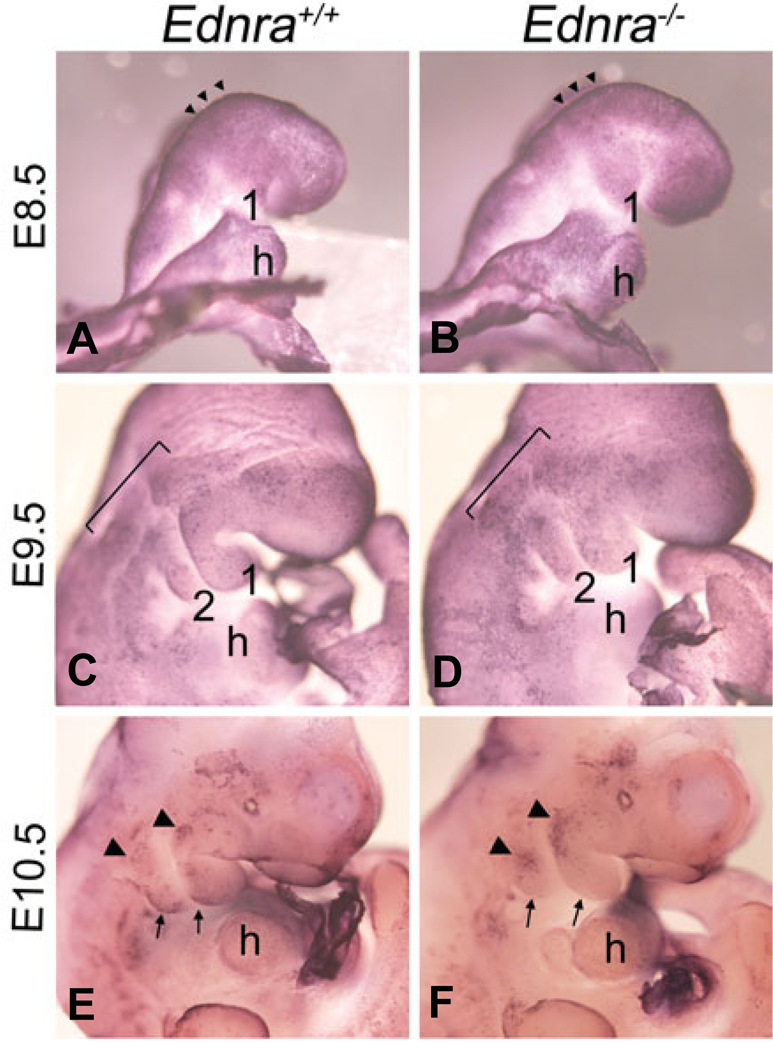
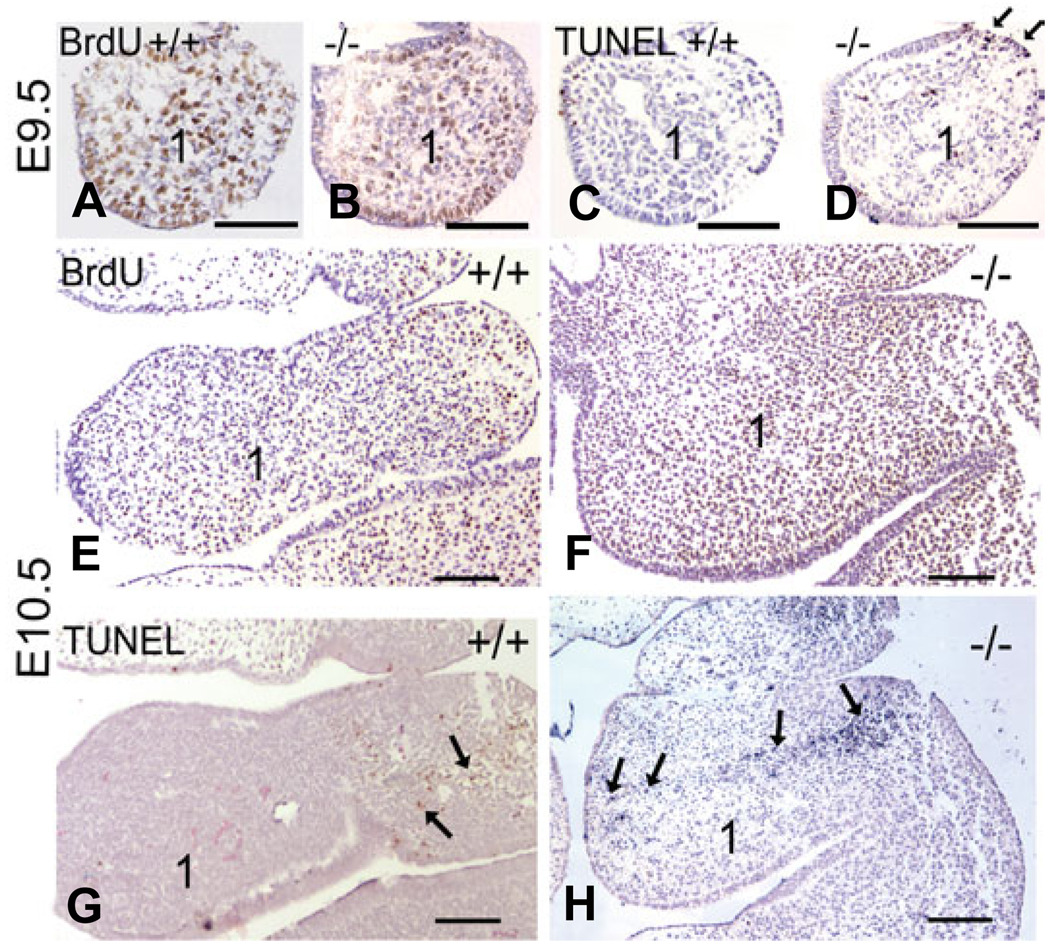
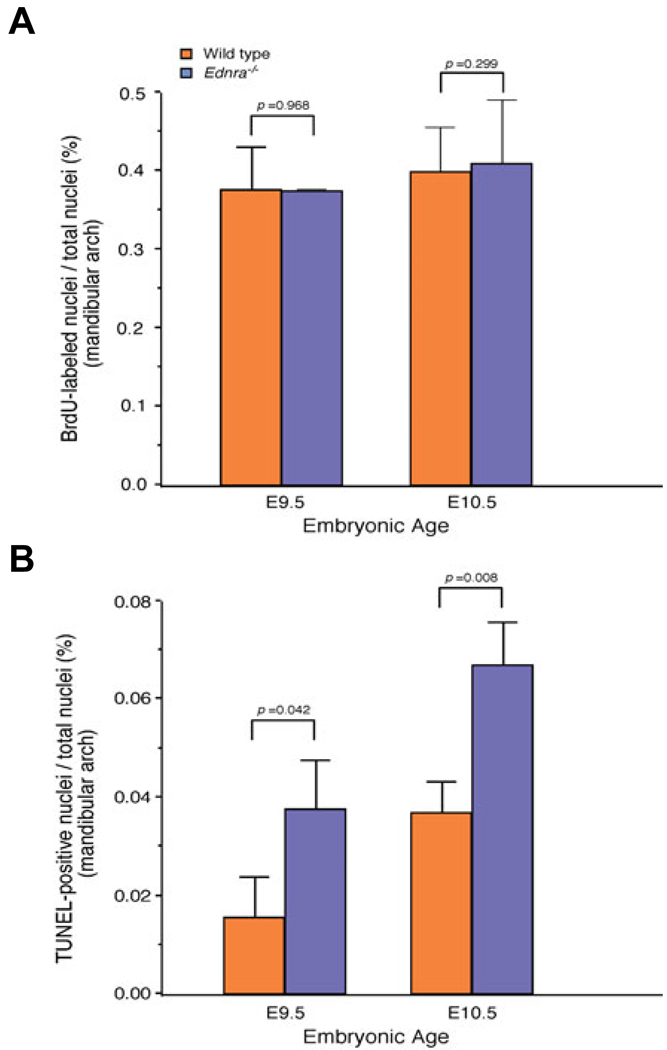
Most of the bone, cartilage and connective tissue of the lower jaw is derived from cranial neural crest cells (NCCs) arising from the posterior midbrain and hindbrain. Multiple factors direct the patterning of these NCCs, including endothelin-1-mediated endothelin A receptor (Edn1/Ednra) signaling. Loss of Ednra signaling results in multiple defects in lower jaw and neck structures, including homeotic transformation of lower jaw structures into upper jaw-like structures. However, since the Ednra gene is expressed by both migrating and post-migrating NCCs, the actual function of Ednra in cranial NCC development is not clear. Ednra signaling could be required for normal migration or guidance of NCCs to the pharyngeal arches or in subsequent events in post-migratory NCCs, including proliferation and survival. To address this question, we performed a fate analysis of cranial NCCs in Ednra-/- embryos using the R26R;Wnt1-Cre reporter system, in which Cre expression within NCCs results in permanent beta-galactosidase activity in NCCs and their derivatives. We find that loss of Ednra does not detectably alter either migration of most cranial NCCs into the mandibular first arch and second arch or their subsequent proliferation. However, mesenchymal cell apoptosis is increased two fold in both E9.5 and E10.5 Ednra-/- embryos, with apoptotic cells being present in and just proximal to the pharyngeal arches. Based on these studies, Ednra signaling appears to be required by most cranial NCCs after they reach the pharyngeal arches. However, a subset of NCCs appear to require Ednra signaling earlier, with loss of Ednra signaling likely leading to premature cessation of migration into or within the arches and subsequent cell death.
Figures

References
-
- Bronner-Fraser M. Origins and developmental potential of the neural crest. Exp. Cell Res. 1995;218:405–417. - PubMed
-
- Chai Y, Jiang X, Ito Y, Bringas P, Han J, Rositch DH, Soriano P, Mcmahon AP, Sucov HM. Fate of the mammalian cranial neural crest during tooth and mandibular morphogenesis. Development. 2000;127:1671–1679. - PubMed
-
- Clouthier DE, Hosoda K, Richardson JA, Williams SC, Yanagisawa H, Kuwaki T, Kumada M, Hammer RE, Yanagisawa M. Cranial and cardiac neural crest defects in endothelin-A receptor-deficient mice. Development. 1998;125:813–824. - PubMed
-
- Clouthier DE, Schilling TF. Understanding endothelin-1 function during craniofacial development in the mouse and zebrafish. Birth Defects Res. (Part C) 2004;72:190–199. - PubMed
-
- Clouthier DE, Williams SC, Hammer RE, Richardson JA, Yanagisawa H. Cell-autonomous and non-autonomous actions of endothelin-A receptor signaling in craniofacial and cardiovascular development. Dev. Biol. 2003;261:506–519. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
