

Physiological osmotic induction of Leptospira interrogans adhesion: LigA and LigB bind extracellular matrix proteins and fibrinogen
- PMID: 17296754
- PMCID: PMC1865782
- DOI: 10.1128/IAI.01635-06
Physiological osmotic induction of Leptospira interrogans adhesion: LigA and LigB bind extracellular matrix proteins and fibrinogen
Abstract
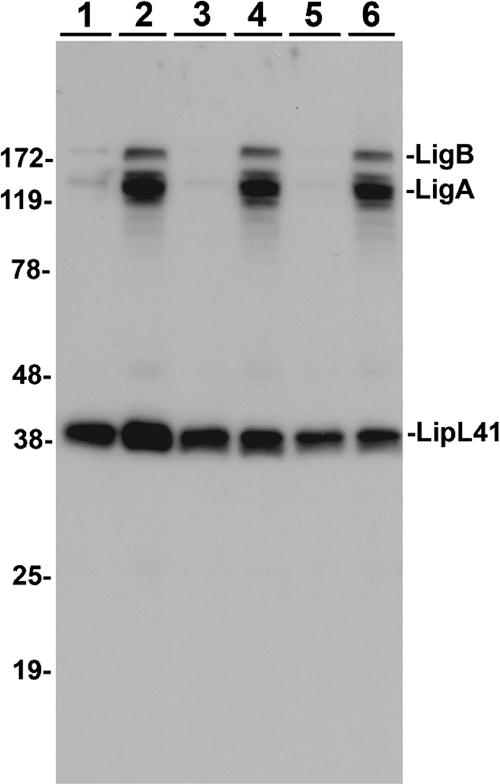
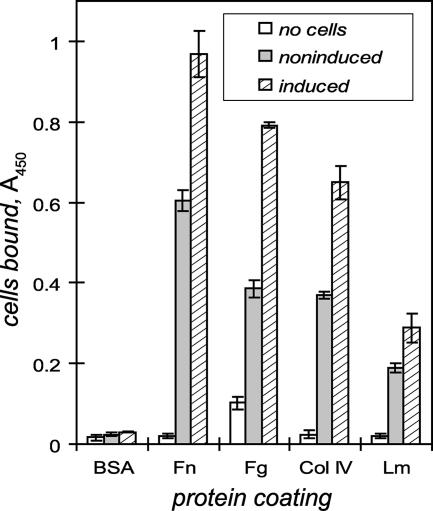
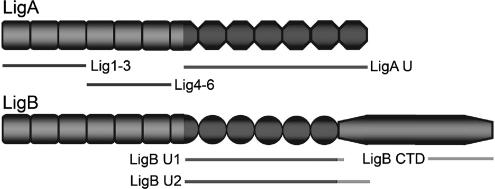
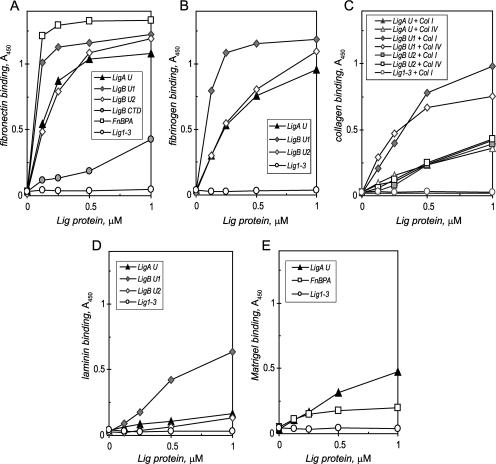
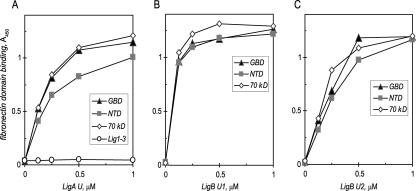
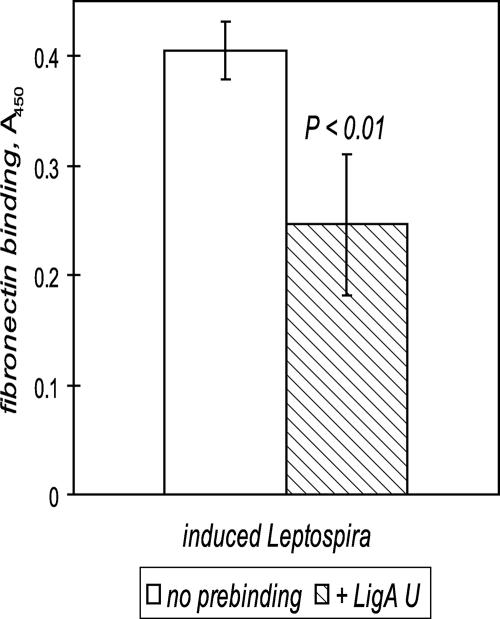
Transmission of leptospirosis occurs through contact of mucous membranes and abraded skin with freshwater contaminated by pathogenic Leptospira spp. Exposure to physiological osmolarity induces leptospires to express high levels of the Lig surface proteins containing imperfect immunoglobulin-like repeats that are shared or differ between LigA and LigB. We report that osmotic induction of Lig is accompanied by 1.6- to 2.5-fold increases in leptospiral adhesion to immobilized extracellular matrix and plasma proteins, including collagens I and IV, laminin, and especially fibronectin and fibrinogen. Recombinant LigA-unique and LigB-unique repeat proteins bind to these same host ligands. We found that the avidity of LigB in binding fibronectin is comparable to that of the Staphylococcus aureus FnBPA D repeats. Both LigA- and LigB-unique repeats interact with the amino-terminal fibrin- and gelatin-binding domains of fibronectin, which are also recognized by fibronectin-binding proteins mediating the adhesion of other microbial pathogens. In contrast, repeats common to both LigA and LigB do not bind these host proteins, and nonrepeat sequences in the carboxy-terminal domain of LigB show only weak interaction with fibronectin and fibrinogen. A functional role for the binding activity of LigA and LigB is suggested by the ability of the recombinants to inhibit leptospiral adhesion to fibronectin by 28% and 21%, respectively. The binding of LigA and LigB to multiple ligands present in different tissues suggests that these adhesins may be involved in the initial colonization and dissemination stages of leptospirosis. The characterization of the Lig adhesin function should aid the design of Lig-based vaccines and serodiagnostic tests.
Figures
References
-
- Ballard, S. A., M. Williamson, B. Adler, T. Vinh, and S. Faine. 1986. Interactions of virulent and avirulent leptospires with primary cultures of renal epithelial cells. J. Med. Microbiol. 21:59-67. - PubMed
-
- Bharti, A. R., J. E. Nally, J. N. Ricaldi, M. A. Matthias, M. M. Diaz, M. A. Lovett, P. N. Levett, R. H. Gilman, M. R. Willig, E. Gotuzzo, and J. M. Vinetz. 2003. Leptospirosis: a zoonotic disease of global importance. Lancet Infect. Dis. 3:757-771. - PubMed
-
- Brown, E. L., J. H. Kim, E. S. Reisenbichler, and M. Höök. 2005. Multicomponent Lyme vaccine: three is not a crowd. Vaccine 23:3687-3696. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
