

Prion species barrier between the closely related yeast proteins is detected despite coaggregation
- PMID: 17296932
- PMCID: PMC1815260
- DOI: 10.1073/pnas.0611158104
Prion species barrier between the closely related yeast proteins is detected despite coaggregation
Abstract
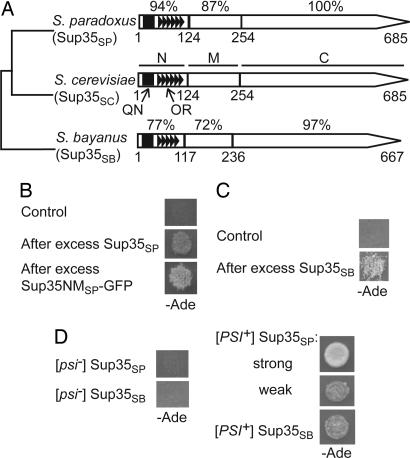
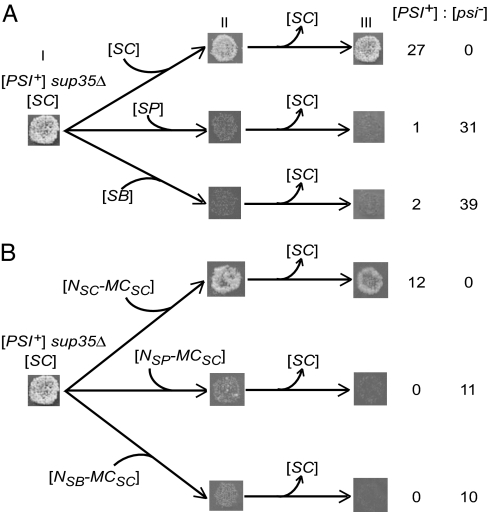
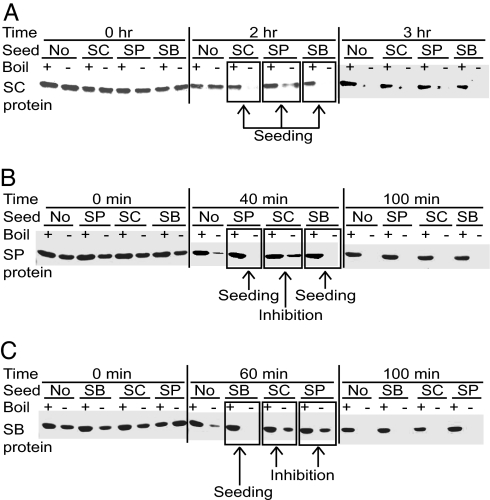
Prions are self-perpetuating and, in most cases, aggregation-prone protein isoforms that transmit neurodegenerative diseases in mammals and control heritable traits in yeast. Prion conversion requires a very high level of identity of the interacting protein sequences. Decreased transmission of the prion state between divergent proteins is termed "species barrier" and was thought to occur because of the inability of divergent prion proteins to coaggregate. Species barrier can be overcome in cross-species infections, e.g., from "mad cows" to humans. We studied the counterparts of yeast prion protein Sup35, originated from three different species of the Saccharomyces sensu stricto group and exhibiting the range of prion domain divergence that overlaps with the range of divergence observed among distant mammalian species. All three proteins were capable of forming a prion in Saccharomyces cerevisiae, although prions formed by heterologous proteins were usually less stable than the endogenous S. cerevisiae prion. Heterologous Sup35 proteins coaggregated in the S. cerevisiae cells. However, in vivo cross-species prion conversion was decreased and in vitro polymerization was cross-inhibited in at least some heterologous combinations, thus demonstrating the existence of prion species barrier. Moreover, the barrier between the S. cerevisiae protein and its Saccharomyces paradoxus and Saccharomyces bayanus counterparts was asymmetric both in vivo and in vitro. Our data show that a decreased cross-species prion transmission does not necessarily correlate with a lack of cross-species coaggregation, suggesting that species-specificity of prion transmission is controlled at the level of conformational transition rather than coaggregation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
