

The Saccharomyces cerevisiae 14-3-3 proteins Bmh1 and Bmh2 directly influence the DNA damage-dependent functions of Rad53
- PMID: 17299042
- PMCID: PMC1797148
- DOI: 10.1073/pnas.0611259104
The Saccharomyces cerevisiae 14-3-3 proteins Bmh1 and Bmh2 directly influence the DNA damage-dependent functions of Rad53
Abstract
In this study, we mutated autophosphorylation sites in Rad53 based on their conservation with previously identified autophosphorylation sites in the mammalian Rad53 ortholog, Chk2. As with wild-type Rad53, the autophosphorylation mutant, rad53-TA, undergoes Mec1/Tel1-dependent interactions with Rad9 and Dun1 in response to genotoxic stress. Whereas rad53-TA in vitro kinase activity is severely impaired, the rad53-TA strains are not completely deficient for cell-cycle checkpoint functions, indicating that the mutant kinase retains a basal level of function. We describe a genetic interaction among Rad53, Dun1, and the 14-3-3 proteins Bmh1 and Bmh2 and present evidence that 14-3-3 proteins directly facilitate Rad53 function in vivo. The data presented account for the previously observed checkpoint defects associated with 14-3-3 mutants in Saccharomyces pombe and Saccharomyces cerevisiae. The 14-3-3 functional interaction appears to modulate Rad53 activity, reminiscent of 14-3-3's effect on human Raf1 kinase and distinct from the indirect mode of regulation by 14-3-3 observed for Chk1 or Cdc25.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
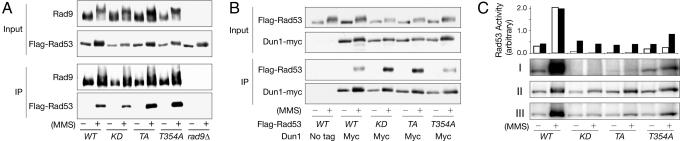


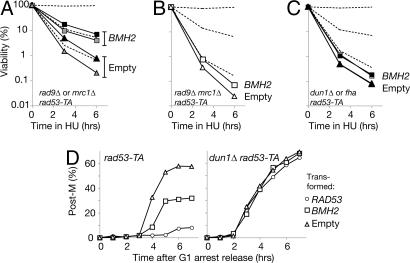


References
-
- Zhou BB, Elledge SJ. Nature. 2000;408:433–439. - PubMed
-
- Nyberg KA, Michelson RJ, Putnam CW, Weinert TA. Annu Rev Genet. 2002;36:617–656. - PubMed
-
- Durocher D, Henckel J, Fersht AR, Jackson SP. Mol Cell. 1999;4:387–394. - PubMed
-
- Manke IA, Lowery DM, Nguyen A, Yaffe MB. Science. 2003;302:636–639. - PubMed
-
- Sanchez Y, Desany BA, Jones WJ, Liu Q, Wang B, Elledge SJ. Science. 1996;271:357–360. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
