

Review
doi: 10.1038/nrn2075.
Epub 2007 Feb 14.
Cell adhesion molecules: signalling functions at the synapse
Affiliations
- PMID: 17299456
- PMCID: PMC4756920
- DOI: 10.1038/nrn2075
Item in Clipboard
Review
Cell adhesion molecules: signalling functions at the synapse
Nat Rev Neurosci.
2007 Mar.
Abstract
Many cell adhesion molecules are localized at synaptic sites in neuronal axons and dendrites. These molecules bridge pre- and postsynaptic specializations but do far more than simply provide a mechanical link between cells. In this review, we will discuss the roles these proteins have during development and at mature synapses. Synaptic adhesion proteins participate in the formation, maturation, function and plasticity of synaptic connections. Together with conventional synaptic transmission mechanisms, these molecules are an important element in the trans-cellular communication mediated by synapses.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
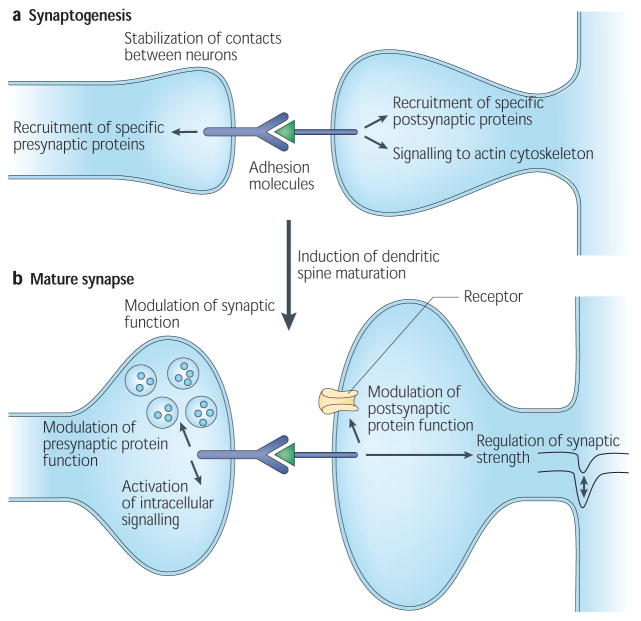
a | At the nascent synaptic site, synaptic adhesion molecules stabilize the initial contact between axons and dendrites. Clustering and binding of adhesion proteins can lead to the recruitment of synaptic proteins via specific cytoplasmic or extracellular domains on these molecules, including PDZ-binding domains. Interactions between adhesion molecules can also lead to the activation of intracellular signalling events that can drive synapse maturation. In particular, signalling to the actin cytoskeleton can lead to the induction of dendritic spine formation. b | In the mature or maturing synapse, synaptic adhesion molecules can interact with channels and other synaptic proteins to modulate their function, either by direct interaction with these proteins or through the activation of intracellular signalling events. In addition, synaptic adhesion proteins can regulate synaptic plasticity.
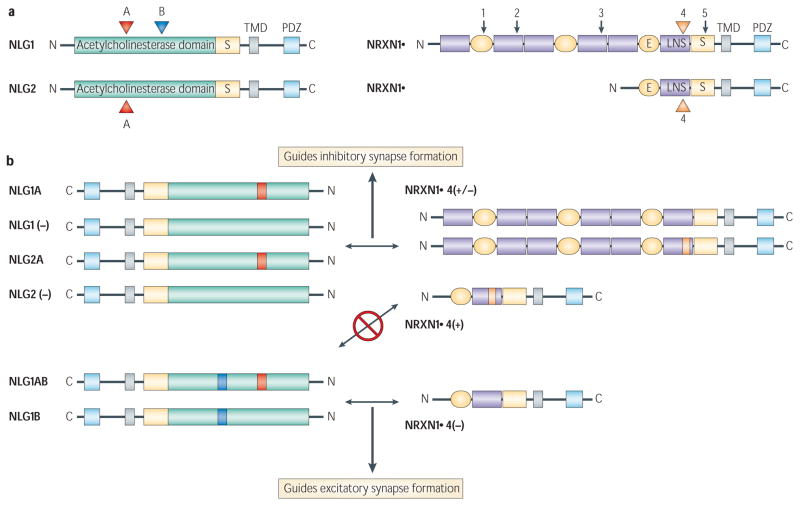
Alternative splicing of presynaptic neurexins and postsynaptic neuroligins can guide the formation of GABA (γ-aminobutyric acid)-containing or glutamatergic synapses. a | Potential sites of inclusion of short alternative exons in neuroligins (NLG) 1 and 2, and neurexin (NRXN) 1α and 1β are shown by arrows and coloured arrowheads. We focus on splice site 4 in neurexins because the most evidence exists for the role of that site in regulating synapse development. b | Shows the splice code that guides interactions between neurexins and neuroligins, as well as formation of excitatory or inhibitory synapses,,. β-neurexins containing the insert at site 4 (orange) or α-neurexins with or without that insert bind neuroligins 1 or 2 with no inserts or containing the insert at site A only (red). These interactions preferentially guide the formation of an inhibitory synaptic contact. Neuroligin 1 containing the insert in site B (blue), with or without the insert in A, is largely restricted to interactions withβ-neurexins lacking an insert at site 4. These interactions preferentially guide the formation of an excitatory synaptic contact. Although these splice variants significantly affect the interactions between neurexins and neuroligins, and formation of excitatory or inhibitory contacts, none is absolute. For example, NLG1(−), NLG1A, NLG2(−), NLG2A and NRXN1β4(−) are more promiscuous and can induce excitatory or inhibitory synapse formation in a reduced in vitro system. In neurons, the function of some of these variants might be restricted by their localization. C, carboxy-terminal domain (cytoplasmic); E, epidermal growth factor-like sequence; LNS, laminin-/neurexin-/sex hormone-binding globulin domain; N, amino-terminal domain (extracellular); PDZ, PDZ binding domain; S, carbohydrate attachment site; TMD, transmembrane domain. Panel a modified, with permission, from REFS , © (2006) Elsevier Science.
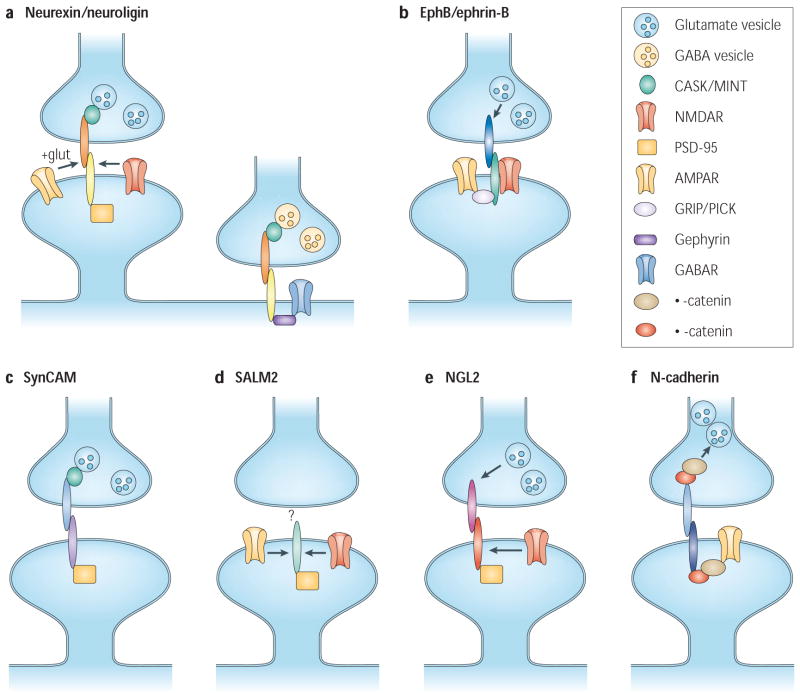
A number of trans-synaptic adhesion molecules are able to control different aspects of synapse development in neuronal cultures and heterologous cell co-cultures. a | Neurexins and neuroligins can induce the formation of both excitatory and inhibitory synapses.b | EphBs can organize both pre- and postsynaptic glutamatergic terminals through mechanisms requiring defined EphB2 protein domains, and signal to induce dendritic spine formation. c | Synaptic cell adhesion molecule (SynCAM) triggers presynaptic maturation but does not yet have a defined role in postsynaptic differentiation. d | Synaptic adhesion-like molecule 2 (SALM2) can regulate the organization of the postsynaptic terminal but not the presynaptic terminal (its presynaptic ligand is unknown). AMPARs (α-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid receptors), and to a lesser degree NMDARs (N-methyl-D-aspartate receptors), can be found at SALM2-induced synapses. e | Netrin G2 ligand (NGL2) can organize pre- and postsynaptic terminals of excitatory synapses. NMDARs are recruited to these sites, but not AMPARs.f | Finally, N-cadherin and catenins are required for the formation of normal presynaptic vesicle reserve pools and have a well-described role in the formation, development and stability of dendritic spines by signalling through Rho GTPases. Cadherins also associate with AMPARs through β-catenins. Arrows indicate that recruitment/clustering occurs, but the mechanism is unclear. ‘?’ indicates that the presynaptic ligand is unknown. CASK, calcium/calmodulin-dependent serine protein kinase; GABAR, γ-aminobutyric acid receptor; GRIP, glutamate receptor interacting protein; MINT, (Munc 18 interacting protein; lin-10/X11); PICK, protein interacting with C kinase; PSD-95, postsynaptic density protein-95.
References
-
- Li Z, Sheng M. Some assembly required: the development of neuronal synapses. Nature Rev Mol Cell Biol. 2003;4:833–841. - PubMed
-
- Waites CL, Craig AM, Garner CC. Mechanisms of vertebrate synaptogenesis. Annu Rev Neurosci. 2005;28:251–274. - PubMed
-
- Kayser MS, Dalva MB. In: Textbook of Neural Repair and Rehabilitation. Selzer ME, Cohen LG, Gage FH, Clarke S, Duncan PW, editors. Vol. 1. Cambridge Univ. Press; Cambridge: 2006. pp. 346–361.
-
- Nguyen T, Sudhof TC. Binding properties of neuroligin 1 and neurexin 1β reveal function as heterophilic cell adhesion molecules. J Biol Chem. 1997;272:26032–26039. - PubMed
-
- Ichtchenko K, et al. Neuroligin 1: a splice site-specific ligand for β-neurexins. Cell. 1995;81:435–443. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
