

CKS1B, overexpressed in aggressive disease, regulates multiple myeloma growth and survival through SKP2- and p27Kip1-dependent and -independent mechanisms
- PMID: 17303695
- PMCID: PMC1885527
- DOI: 10.1182/blood-2006-07-038703
CKS1B, overexpressed in aggressive disease, regulates multiple myeloma growth and survival through SKP2- and p27Kip1-dependent and -independent mechanisms
Abstract
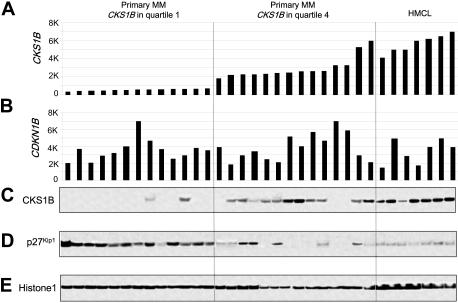
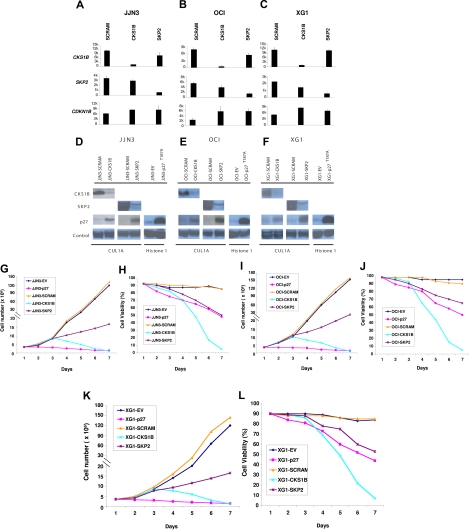
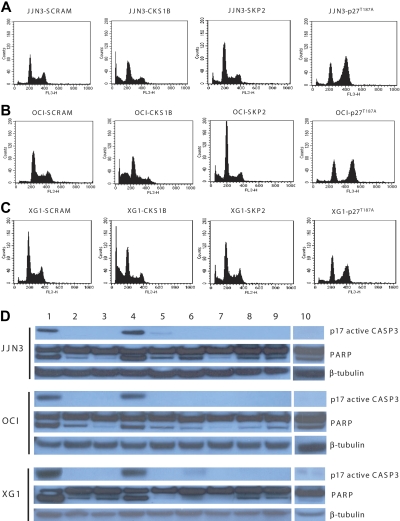
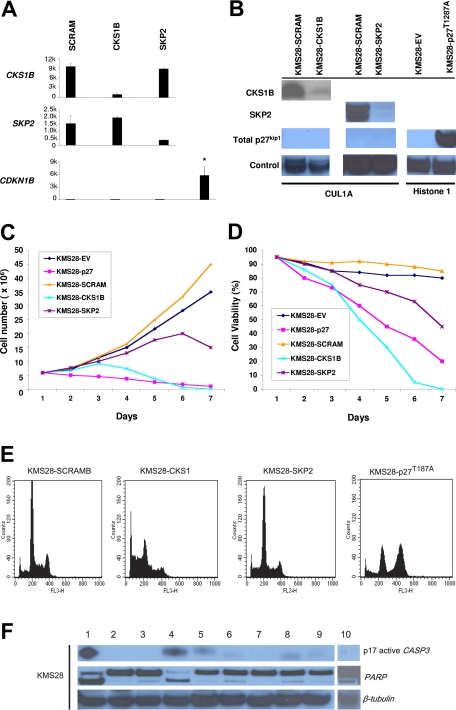
Overexpression of CKS1B, a gene mapping within a minimally amplified region between 153 to 154 Mb of chromosome 1q21, is linked to a poor prognosis in multiple myeloma (MM). CKS1B binds to and activates cyclin-dependent kinases and also interacts with SKP2 to promote the ubiquitination and proteasomal degradation of p27(Kip1). Overexpression of CKS1B or SKP2 contributes to increased p27(Kip1) turnover, cell proliferation, and a poor prognosis in many tumor types. Using 4 MM cell lines harboring MAF-, FGFR3/MMSET-, or CCND1-activating translocations, we show that lentiviral delivery of shRNA directed against CKS1B resulted in ablation of CKS1B mRNA and protein with concomitant stabilization of p27(Kip1), cell cycle arrest, and apoptosis. Although shRNA-mediated knockdown of SKP2 and forced expression of a nondegradable form of p27(Kip1) (p27(T187A)) led to cell cycle arrest, apoptosis was modest. Of importance, while knockdown of SKP2 or overexpression of p27(T187A) induced cell cycle arrest in KMS28PE, an MM cell line with biallelic deletion of CDKN1B/p27(Kip1), CKS1B ablation induced strong apoptosis. These data suggest that CKS1B influences myeloma cell growth and survival through SKP2- and p27(Kip1)-dependent and -independent mechanisms and that therapeutic strategies aimed at abolishing CKS1B function may hold promise for the treatment of high-risk disease for which effective therapies are currently lacking.
Figures
Similar articles
-
CKS1B overexpression implicates clinical aggressiveness of hepatocellular carcinomas but not p27(Kip1) protein turnover: an independent prognosticator with potential p27 (Kip1)-independent oncogenic attributes?Ann Surg Oncol. 2010 Mar;17(3):907-22. doi: 10.1245/s10434-009-0779-8. Epub 2009 Oct 29. Ann Surg Oncol. 2010. PMID: 19866239
-
Amplification and overexpression of CKS1B at chromosome band 1q21 is associated with reduced levels of p27Kip1 and an aggressive clinical course in multiple myeloma.Hematology. 2005;10 Suppl 1:117-26. doi: 10.1080/10245330512331390140. Hematology. 2005. PMID: 16188652
-
Over-expression of CKS1B activates both MEK/ERK and JAK/STAT3 signaling pathways and promotes myeloma cell drug-resistance.Oncotarget. 2010 May;1(1):22-33. doi: 10.18632/oncotarget.105. Oncotarget. 2010. PMID: 20930946 Free PMC article.
-
Gene expression profiling defines a high-risk entity of multiple myeloma.Zhong Nan Da Xue Xue Bao Yi Xue Ban. 2007 Apr;32(2):191-203. Zhong Nan Da Xue Xue Bao Yi Xue Ban. 2007. PMID: 17478923 Review.
-
Proliferation unleashed: the role of Skp2 in vascular smooth muscle cell proliferation.Front Biosci (Landmark Ed). 2011 Jan 1;16(4):1517-35. doi: 10.2741/3802. Front Biosci (Landmark Ed). 2011. PMID: 21196245 Review.
Cited by
-
Ixazomib-Thalidomide-Dexamethasone for induction therapy followed by Ixazomib maintenance treatment in patients with relapsed/refractory multiple myeloma.Br J Cancer. 2019 Oct;121(9):751-757. doi: 10.1038/s41416-019-0581-8. Epub 2019 Sep 27. Br J Cancer. 2019. PMID: 31558804 Free PMC article. Clinical Trial.
-
Factors Explaining the Coincidence of Smoldering Multiple Myeloma and Primary Biliary Cholangitis: A Case Report.Cureus. 2022 Jul 13;14(7):e26830. doi: 10.7759/cureus.26830. eCollection 2022 Jul. Cureus. 2022. PMID: 35974869 Free PMC article.
-
ARNT/HIF-1β links high-risk 1q21 gain and microenvironmental hypoxia to drug resistance and poor prognosis in multiple myeloma.Cancer Med. 2018 Aug;7(8):3899-3911. doi: 10.1002/cam4.1596. Epub 2018 Jun 21. Cancer Med. 2018. PMID: 29926531 Free PMC article.
-
Drug resistance in multiple myeloma: latest findings and new concepts on molecular mechanisms.Oncotarget. 2013 Dec;4(12):2186-207. doi: 10.18632/oncotarget.1497. Oncotarget. 2013. PMID: 24327604 Free PMC article. Review.
-
Multi-omic profiling of simultaneous ductal carcinoma in situ and invasive breast cancer.Breast Cancer Res Treat. 2024 Jun;205(3):451-464. doi: 10.1007/s10549-024-07270-5. Epub 2024 Mar 24. Breast Cancer Res Treat. 2024. PMID: 38523186 Free PMC article.
References
-
- Shaughnessy JD, Jr, Zhan F, Burington BE, et al. A validated gene expression model of high-risk multiple myeloma is defined by deregulated expression of genes mapping to chromosome 1. Prepublished on November 14, 2006, as 10.1182/blood-2006-07-038430. (Now available as Blood. 2007;109:2276–2284.) - DOI - PubMed
-
- Hanamura I, Stewart JP, Huang Y, et al. Frequent gain of chromosome band 1q21 in plasma cell dyscrasias detected by fluorescence in situ hybridization: incidence increases from MGUS to relapsed myeloma and is related to prognosis and disease progression following tandem stem cell transplantation. Blood. 2006;108:1724–1732. - PMC - PubMed
-
- Sawyer JR, Tricot G, Mattox S, Jagannath S, Barlogie B. Jumping translocations of chromosome 1q in multiple myeloma: evidence for a mechanism involving decondensation of pericentromeric heterochromatin. Blood. 1998;91:1732–1741. - PubMed
-
- Le Baccon P, Leroux D, Dascalescu C, et al. Novel evidence of a role for chromosome 1 pericentric heterochromatin in the pathogenesis of B-cell lymphoma and multiple myeloma. Genes Chromosomes Cancer. 2001;32:250–264. - PubMed
-
- Sawyer JR, Tricot G, Lukacs JL, et al. Genomic instability in multiple myeloma: evidence for jumping segmental duplications of chromosome arm 1q. Genes Chromosomes Cancer. 2005;42:95–106. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
