

HER-2/neu antigen loss and relapse of mammary carcinoma are actively induced by T cell-mediated anti-tumor immune responses
- PMID: 17304628
- PMCID: PMC3732067
- DOI: 10.1002/eji.200636639
HER-2/neu antigen loss and relapse of mammary carcinoma are actively induced by T cell-mediated anti-tumor immune responses
Abstract
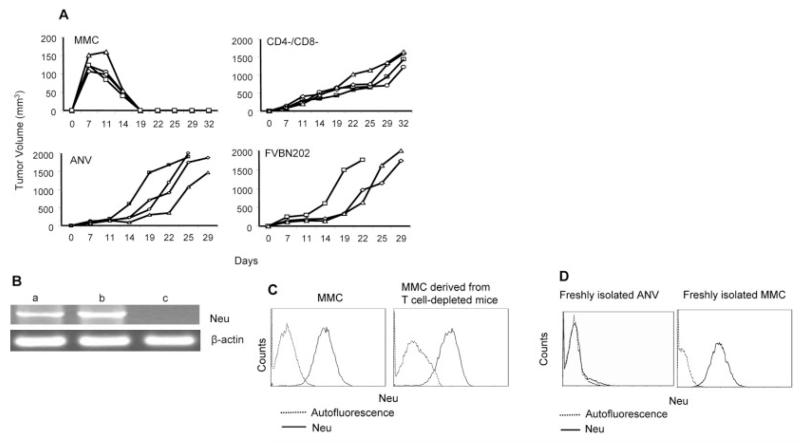
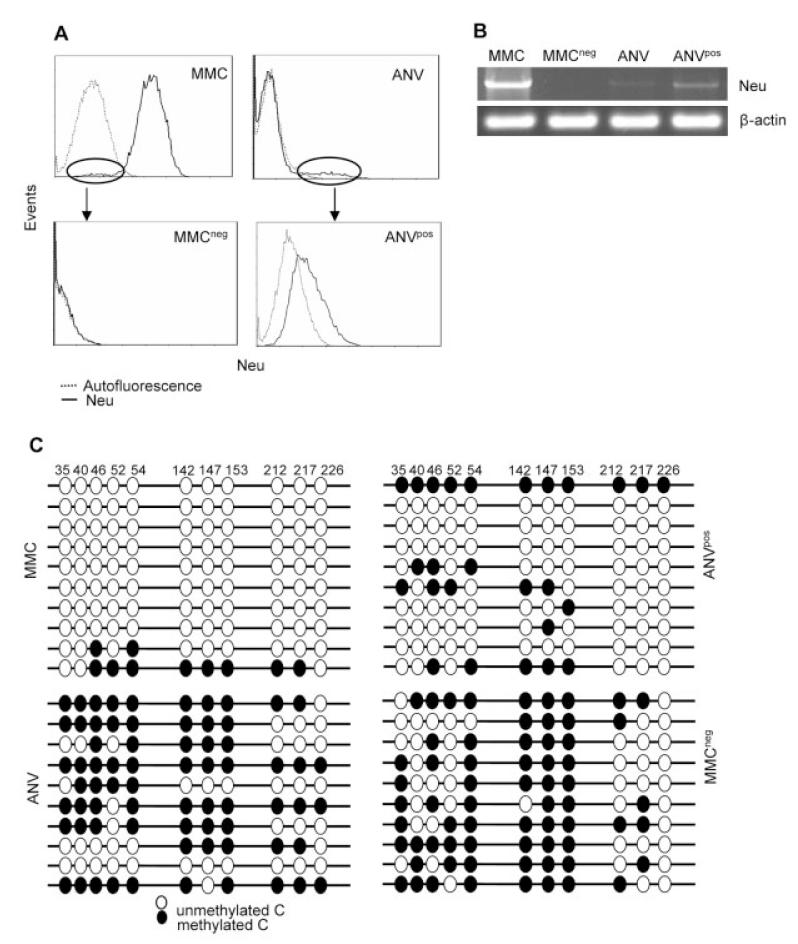
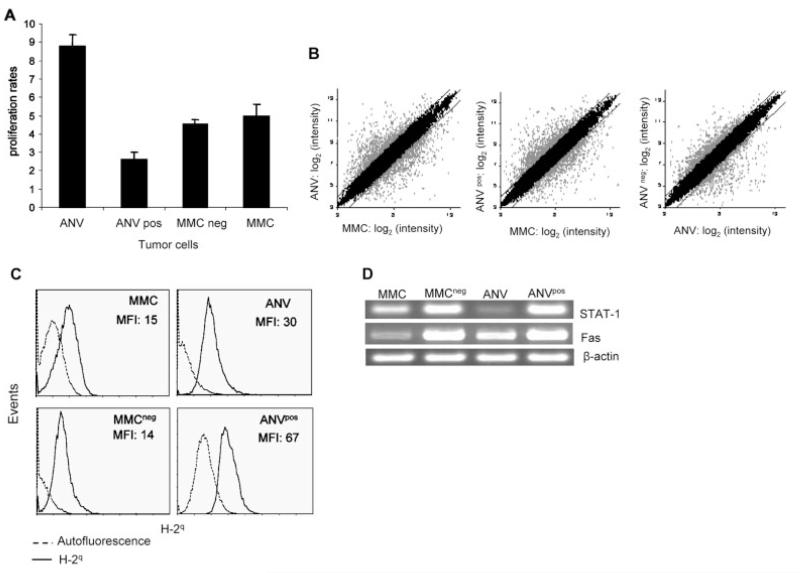
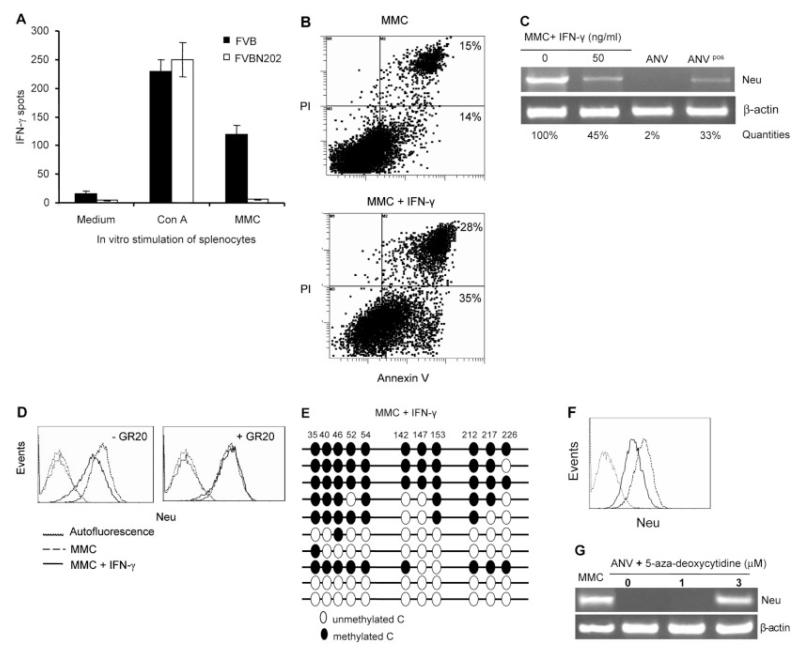
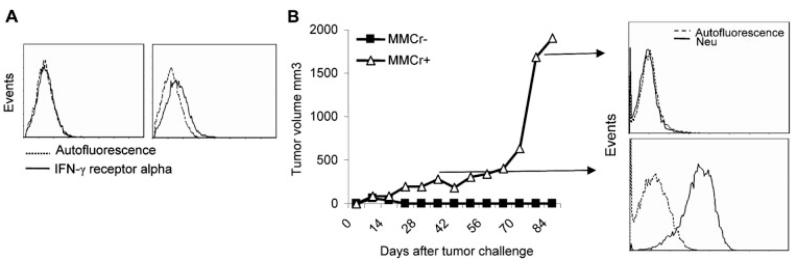
Induction of tumor-specific immune responses results in the inhibition of tumor development. However, tumors recur because of the tumor immunoediting process that facilitates development of escape mechanisms in tumors. It is not known whether tumor escape is an active process whereby anti-tumor immune responses induce loss or downregulation of the target antigen in the antigen-positive clones. To address this question, we used rat neu-overexpressing mouse mammary carcinoma (MMC) and its relapsed neu antigen-negative variant (ANV). ANV emerged from MMC under pressure from neu-specific T cell responses in vivo. We then cloned residual neu antigen-negative cells from MMC and residual neu antigen-positive cells from ANV. We found marked differences between these neu-negative clones and ANV, demonstrating that the residual neu-negative clones are probably not the origin of ANV. Since initial rejection of MMC was associated with the presence of IFN-gamma-secreting T cells, we treated MMC with IFN-gamma and showed that IFN-gamma could induce downregulation of neu expression in MMC. This appears to be due to methylation of the neu promoter. Together, these data suggest that neu antigen loss is an active process that occurs in primary tumors due to the neu-targeted anti-tumor immune responses.
Figures
References
-
- Maeurer MJ, Gollin SM, Storkus WJ, Swaney W, Karbach J, Martin D, Castelli C, et al. Tumor escape from immune recognition: lethal recurrent melanoma in a patient associated with downregulation of the peptide transporter protein TAP-1 and loss of expression of the immunodominant MART-1/Melan-A antigen. J. Clin. Invest. 1996;98:1633–1641. - PMC - PubMed
-
- Yamshchikov GV, Mullins DW, Chang CC, Ogino T, Thompson L, Presley J, Galavotti H, et al. Sequential immune escape and shifting of Tcell responses in a long-term survivor of melanoma. J. Immunol. 2005;174:6863–6871. - PubMed
-
- Facoetti A, Nano R, Zelini P, Morbini P, Benericetti E, Ceroni M, Campoli M, Ferrone S. Human leukocyte antigen and antigen processing machinery component defects in astrocytic tumors. Clin. Cancer Res. 2005;11:8304–8311. - PubMed
-
- Shankaran V, Ikeda H, Bruce AT, White JM, Swanson PE, Old LJ, Schreiber RD. IFN-γ and lymphocytes prevent primary tumour development and shape tumour immunogenicity. Nature. 2001;410:1107–1111. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
