

FoxH1 negatively modulates flk1 gene expression and vascular formation in zebrafish
- PMID: 17306248
- PMCID: PMC1876740
- DOI: 10.1016/j.ydbio.2007.01.023
FoxH1 negatively modulates flk1 gene expression and vascular formation in zebrafish
Abstract
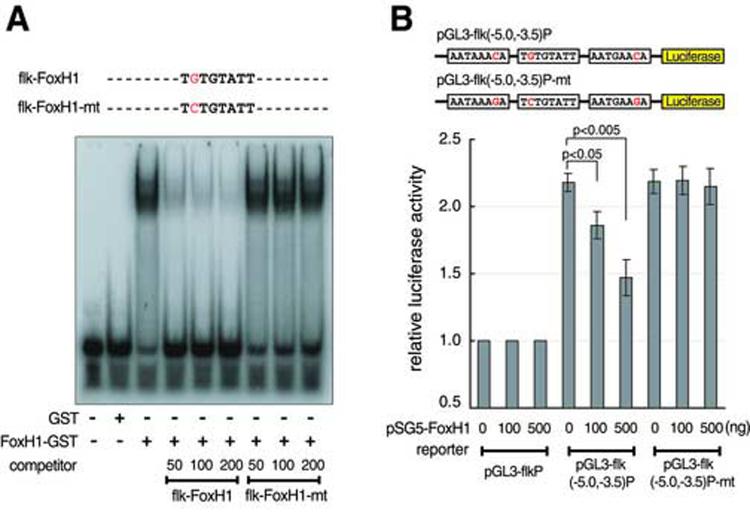
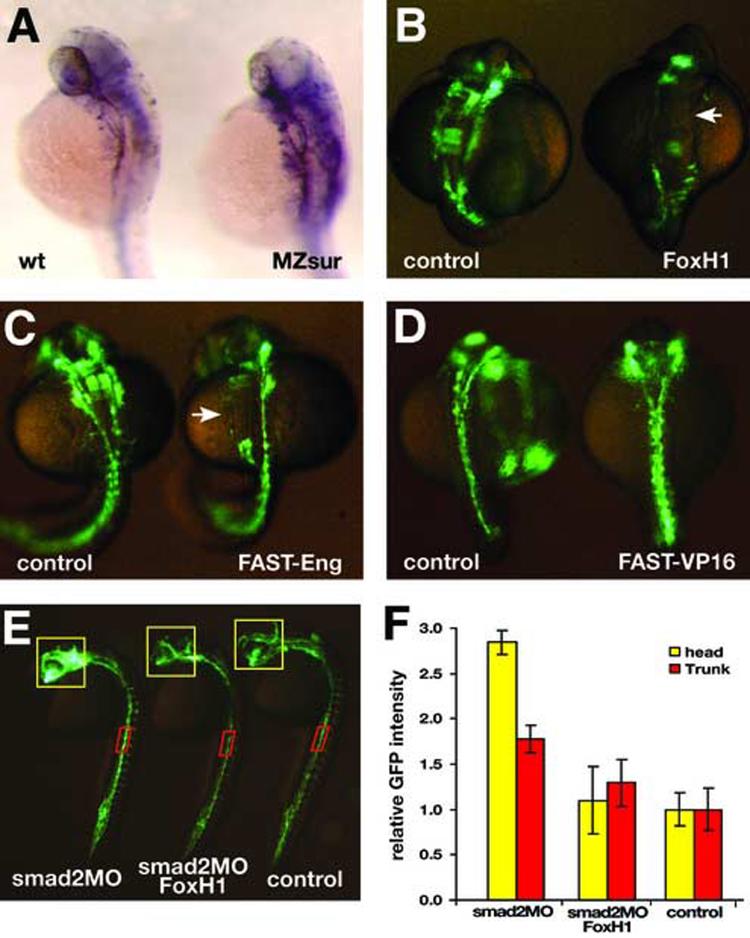
Flk1 is the major receptor for VEGF on endothelial cells. During embryogenesis, flk1 is required for both vasculogenesis and angiogenesis and abnormally elevated flk1 expression is often associated with pathological conditions in adults. While the biological function of flk1 has been studied extensively, very little is known about how the flk1 gene is regulated at the transcriptional level. Our transgenic study led to the identification of a flk1 endothelial enhancer positioned approximately 5 kb upstream of the flk1 translation initiation site. Binding sites for FoxH1, scl, ets and gata factors are found in the zebrafish flk1 endothelial enhancer, as well as in upstream sequences of mouse flk1 and human kdr genes, suggesting that the regulatory machinery for flk1/kdr is conserved from fish to mammals. The roles of scl, ets and gata factors in hemangioblasts have been well defined, but the significance of FoxH1 in vessel formation has not been explored previously. Here we show that FoxH1 binds to the flk1 endothelial enhancer in vitro and functions as a repressor for flk1 transcription in cultured cells. Consistent with these findings, the expression level of flk1 is elevated in embryos lacking both maternal and zygotic FoxH1. We further show that overexpression of FoxH1 has a negative effect on vascular formation that can be counteracted by the down-regulation of smad2 activity in zebrafish embryos. Taken together, our data provide the first evidence that flk1 is a direct target of FoxH1 and that FoxH1 is involved in vessel formation in zebrafish.
Figures
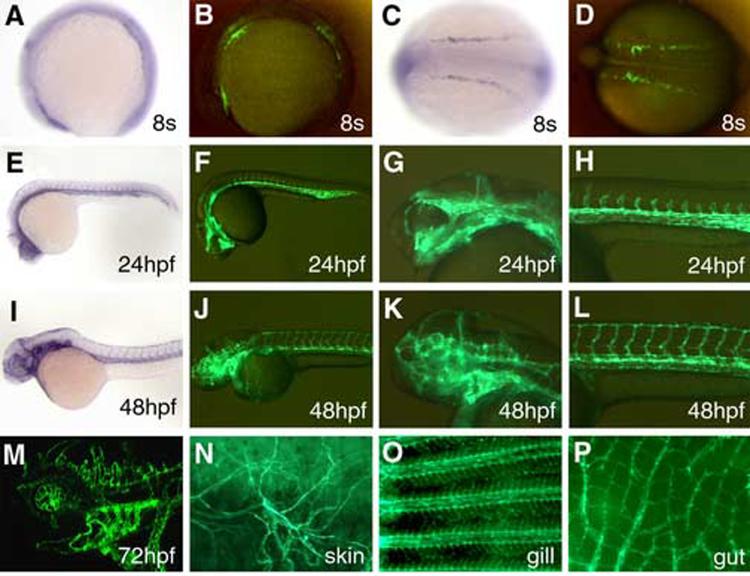
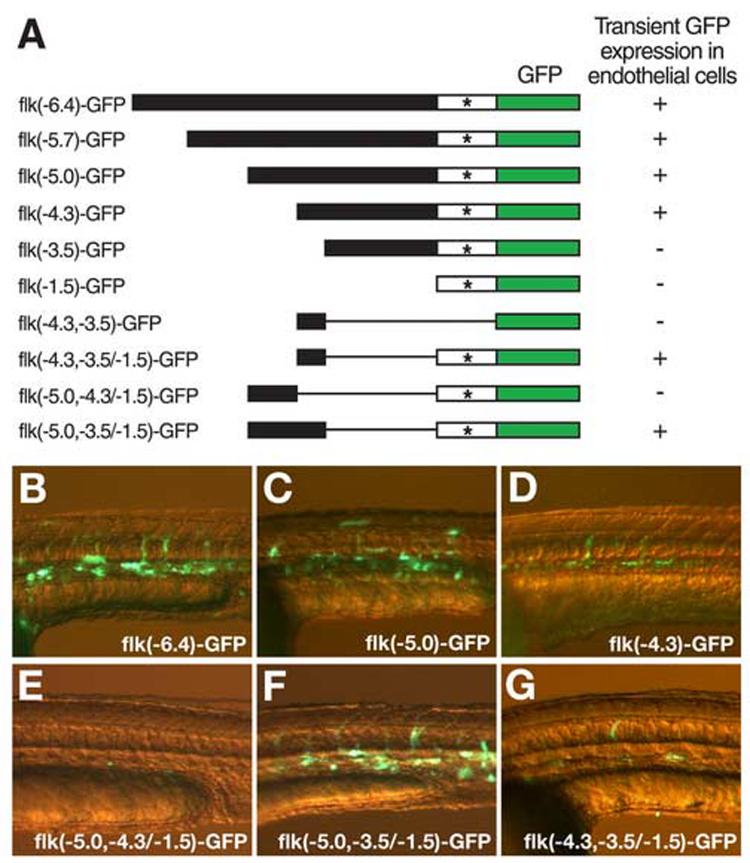
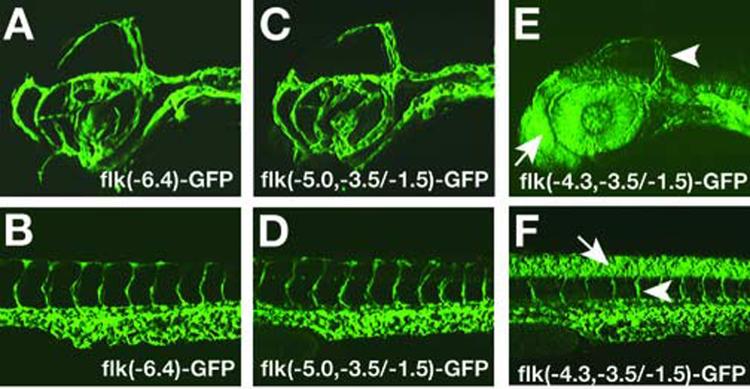
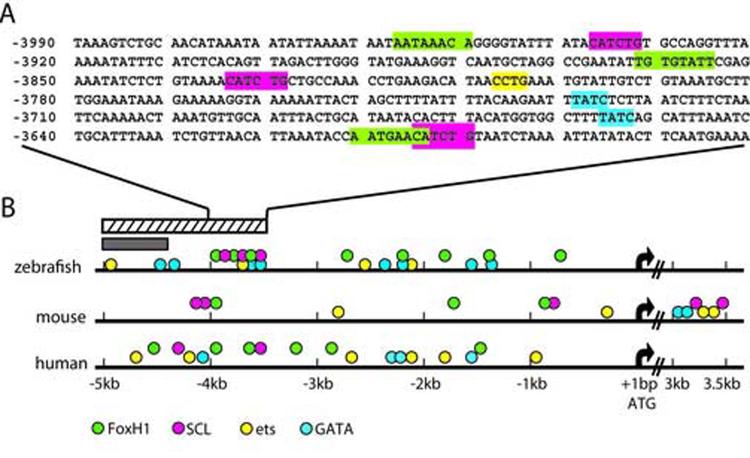
Similar articles
-
An Intronic Flk1 Enhancer Directs Arterial-Specific Expression via RBPJ-Mediated Venous Repression.Arterioscler Thromb Vasc Biol. 2016 Jun;36(6):1209-19. doi: 10.1161/ATVBAHA.116.307517. Epub 2016 Apr 14. Arterioscler Thromb Vasc Biol. 2016. PMID: 27079877 Free PMC article.
-
Analysis of a zebrafish VEGF receptor mutant reveals specific disruption of angiogenesis.Curr Biol. 2002 Aug 20;12(16):1405-12. doi: 10.1016/s0960-9822(02)01044-8. Curr Biol. 2002. PMID: 12194822
-
Posterior-anterior gradient of zebrafish hes6 expression in the presomitic mesoderm is established by the combinatorial functions of the downstream enhancer and 3'UTR.Dev Biol. 2016 Jan 15;409(2):543-54. doi: 10.1016/j.ydbio.2015.11.010. Epub 2015 Nov 17. Dev Biol. 2016. PMID: 26596999
-
ETS Transcription Factor ETV2/ER71/Etsrp in Hematopoietic and Vascular Development.Curr Top Dev Biol. 2016;118:77-111. doi: 10.1016/bs.ctdb.2016.01.005. Epub 2016 Feb 12. Curr Top Dev Biol. 2016. PMID: 27137655 Review.
-
[Vascular endothelial cell development and underlying mechanisms].Yi Chuan. 2012 Sep;34(9):1114-22. Yi Chuan. 2012. PMID: 23017452 Review. Chinese.
Cited by
-
Context-specific interactions between Notch and ALK1 cannot explain ALK1-associated arteriovenous malformations.Cardiovasc Res. 2015 Jul 1;107(1):143-52. doi: 10.1093/cvr/cvv148. Epub 2015 May 12. Cardiovasc Res. 2015. PMID: 25969392 Free PMC article.
-
A limb-girdle muscular dystrophy 2I model of muscular dystrophy identifies corrective drug compounds for dystroglycanopathies.JCI Insight. 2018 Sep 20;3(18):e120493. doi: 10.1172/jci.insight.120493. eCollection 2018 Sep 20. JCI Insight. 2018. PMID: 30232282 Free PMC article.
-
Genome-wide view of TGFβ/Foxh1 regulation of the early mesendoderm program.Development. 2014 Dec;141(23):4537-47. doi: 10.1242/dev.107227. Epub 2014 Oct 30. Development. 2014. PMID: 25359723 Free PMC article.
-
Fine-tune regulation of carboxypeptidase N1 controls vascular patterning during zebrafish development.Sci Rep. 2017 May 12;7(1):1852. doi: 10.1038/s41598-017-01976-x. Sci Rep. 2017. PMID: 28500283 Free PMC article.
-
Nmnat1-Rbp7 Is a Conserved Fusion-Protein That Combines NAD+ Catalysis of Nmnat1 with Subcellular Localization of Rbp7.PLoS One. 2015 Nov 30;10(11):e0143825. doi: 10.1371/journal.pone.0143825. eCollection 2015. PLoS One. 2015. PMID: 26618989 Free PMC article.
References
-
- Attisano L, Silvestri C, Izzi L, Labbe E. The transcriptional role of Smads and FAST (FoxH1) in TGFbeta and activin signalling. Mol Cell Endocrinol. 2001;180:3–11. - PubMed
-
- Beis D, Bartman T, Jin SW, Scott IC, D'Amico LA, Ober EA, Verkade H, Frantsve J, Field HA, Wehman A, Baier H, Tallafuss A, Bally-Cuif L, Chen JN, Stainier DY, Jungblut B. Genetic and cellular analyses of zebrafish atrioventricular cushion and valve development. Development. 2005;132:4193–204. - PubMed
-
- Chen G, Nomura M, Morinaga H, Matsubara E, Okabe T, Goto K, Yanase T, Zheng H, Lu J, Nawata H. Modulation of androgen receptor transactivation by FoxH1. A newly identified androgen receptor corepressor. J Biol Chem. 2005;280:36355–63. - PubMed
-
- Chen X, Rubock MJ, Whitman M. A transcriptional partner for MAD proteins in TGF-beta signalling. Nature. 1996;383:691–6. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
