

Functional genomic analysis supports conservation of function among cellulose synthase-like a gene family members and suggests diverse roles of mannans in plants
- PMID: 17307900
- PMCID: PMC1851810
- DOI: 10.1104/pp.106.093989
Functional genomic analysis supports conservation of function among cellulose synthase-like a gene family members and suggests diverse roles of mannans in plants
Abstract
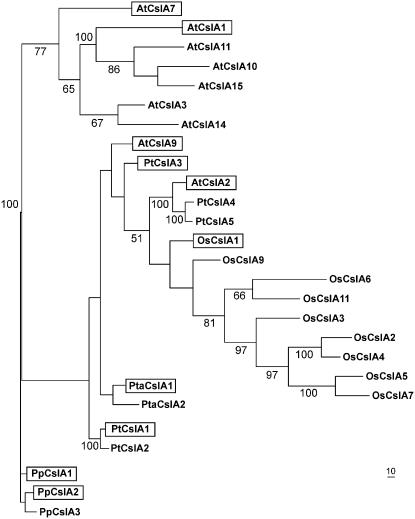
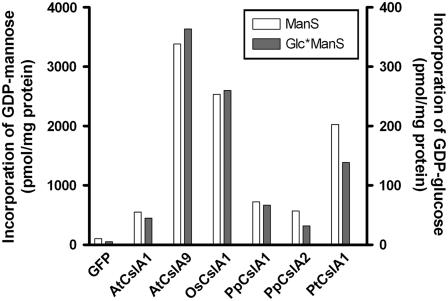
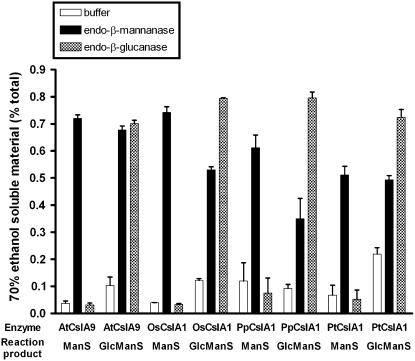
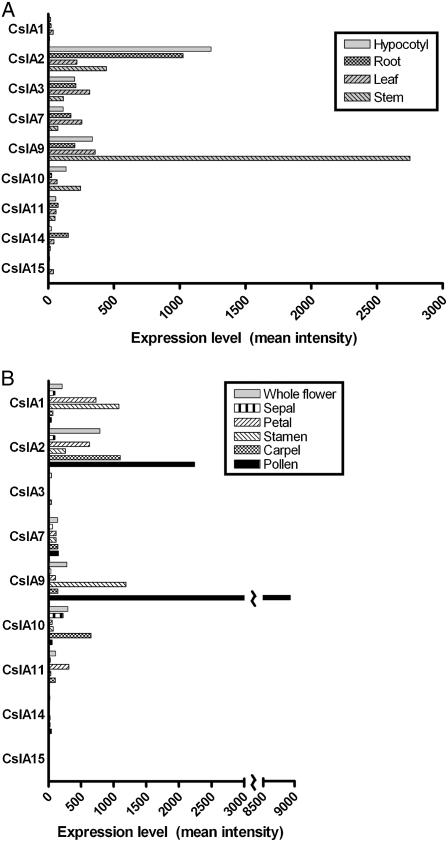
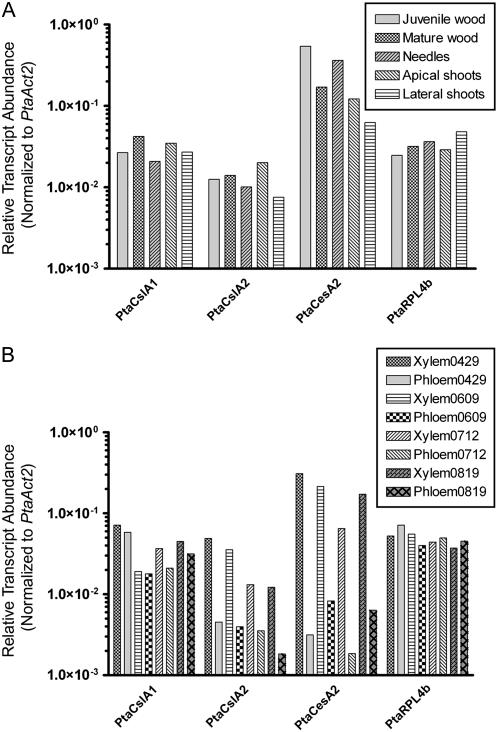
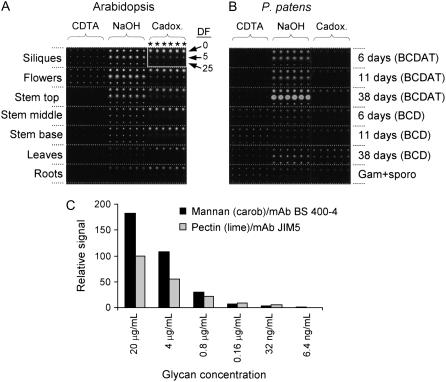
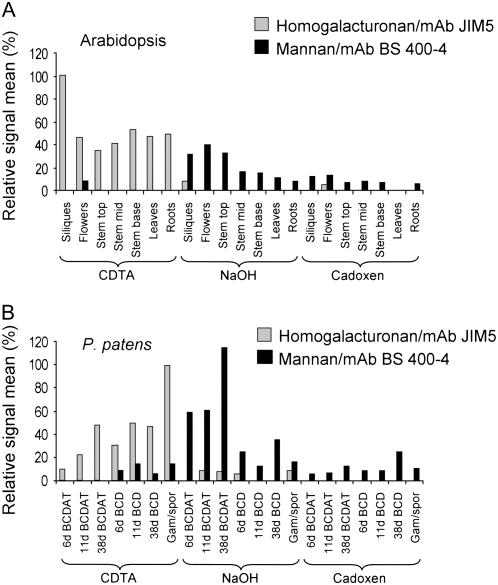
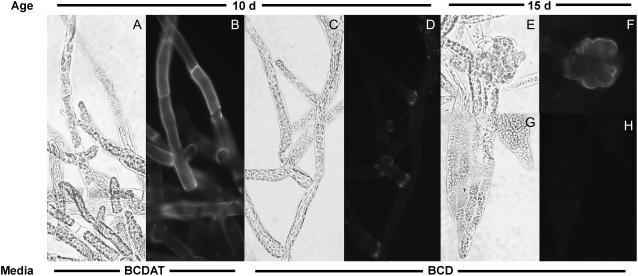
Mannan polysaccharides are widespread among plants, where they serve as structural elements in cell walls, as carbohydrate reserves, and potentially perform other important functions. Previous work has demonstrated that members of the cellulose synthase-like A (CslA) family of glycosyltransferases from Arabidopsis (Arabidopsis thaliana), guar (Cyamopsis tetragonolobus), and Populus trichocarpa catalyze beta-1,4-mannan and glucomannan synthase reactions in vitro. Mannan polysaccharides and homologs of CslA genes appear to be present in all lineages of land plants analyzed to date. In many plants, the CslA genes are members of extended multigene families; however, it is not known whether all CslA proteins are glucomannan synthases. CslA proteins from diverse land plant species, including representatives of the mono- and dicotyledonous angiosperms, gymnosperms, and bryophytes, were produced in insect cells, and each CslA protein catalyzed mannan and glucomannan synthase reactions in vitro. Microarray mining and quantitative real-time reverse transcription-polymerase chain reaction analysis demonstrated that transcripts of Arabidopsis and loblolly pine (Pinus taeda) CslA genes display tissue-specific expression patterns in vegetative and floral tissues. Glycan microarray analysis of Arabidopsis indicated that mannans are present throughout the plant and are especially abundant in flowers, siliques, and stems. Mannans are also present in chloronemal and caulonemal filaments of Physcomitrella patens, where they are prevalent at cell junctions and in buds. Taken together, these results demonstrate that members of the CslA gene family from diverse plant species encode glucomannan synthases and support the hypothesis that mannans function in metabolic networks devoted to other cellular processes in addition to cell wall structure and carbohydrate storage.
Figures
References
-
- Auxtova O, Liskova D, Kakoniova D, Kubackova M, Karacsonyi S, Bilisics L (1995) Effect of galactoglucomannan-derived oligosaccharides on elongation growth of pea and spruce stem segments stimulated by auxin. Planta 196 420–424
-
- Baldauf SL (2003) Phylogeny for the faint of heart: a tutorial. Trends Genet 19 345–351 - PubMed
-
- Benova-Kakosova A, Digonnet C, Goubet F, Ranocha P, Jauneau A, Pesquet E, Barbier O, Zhang Z, Capek P, Dupree P, et al (2006) Galactoglucomannans increase cell population density and alter the protoxylem/metaxylem tracheary element ratio in xylogenic cultures of zinnia. Plant Physiol 142 696–709 - PMC - PubMed
-
- Bilisics L, Vojtassak J, Capek P, Kollarova K, Liskova D (2004) Changes in glycosidase activities during galactoglucomannan oligosaccharide inhibition of auxin induced growth. Phytochemistry 65 1903–1909 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
