

Interaction between trichosanthin, a ribosome-inactivating protein, and the ribosomal stalk protein P2 by chemical shift perturbation and mutagenesis analyses
- PMID: 17308345
- PMCID: PMC1865052
- DOI: 10.1093/nar/gkm065
Interaction between trichosanthin, a ribosome-inactivating protein, and the ribosomal stalk protein P2 by chemical shift perturbation and mutagenesis analyses
Abstract
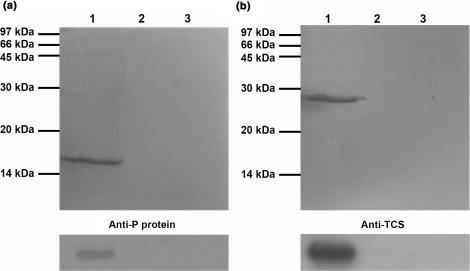
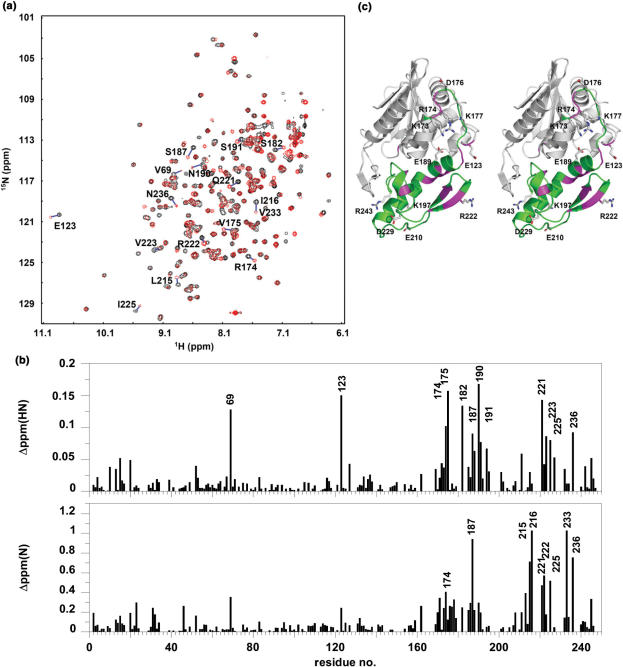
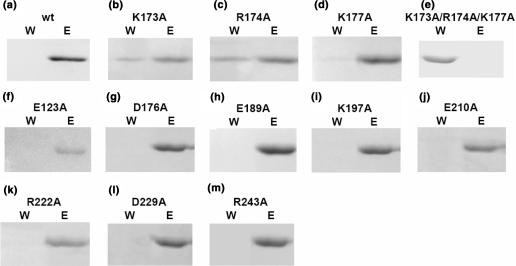
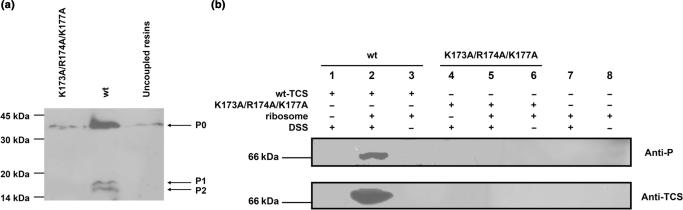
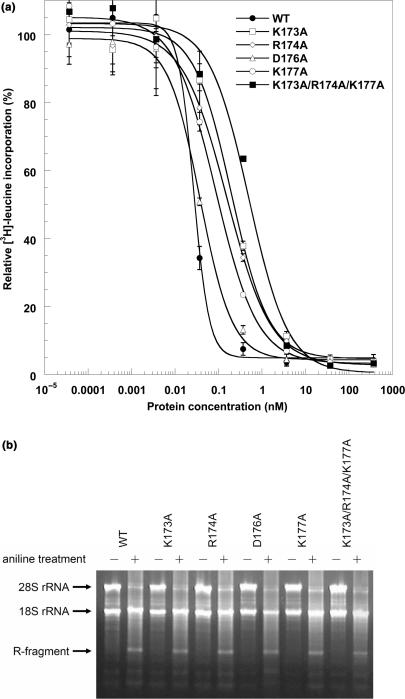
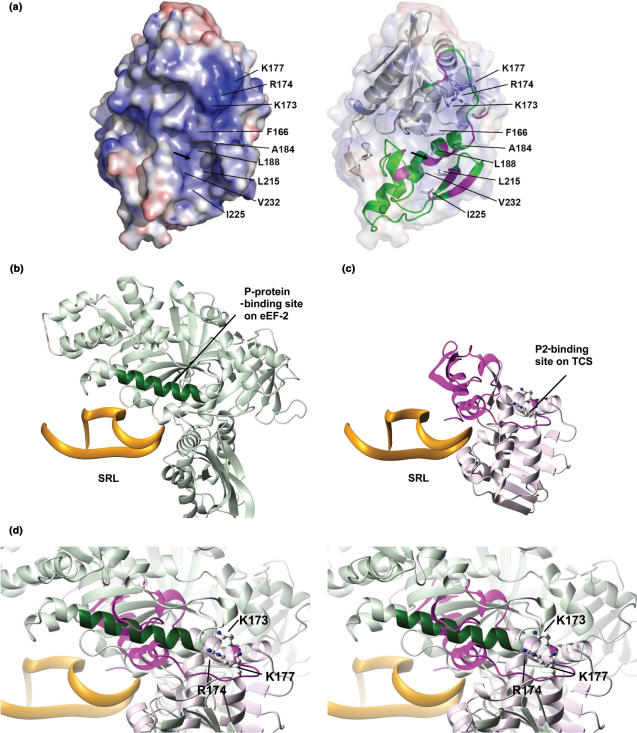
Trichosanthin (TCS) is a type I ribosome-inactivating protein that inactivates ribosome by enzymatically depurinating the A(4324) at the alpha-sarcin/ricin loop of 28S rRNA. We have shown in this and previous studies that TCS interacts with human acidic ribosomal proteins P0, P1 and P2, which constitute the lateral stalk of eukaryotic ribosome. Deletion mutagenesis showed that TCS interacts with the C-terminal tail of P2, the sequences of which are conserved in P0, P1 and P2. The P2-binding site on TCS was mapped to the C-terminal domain by chemical shift perturbation experiments. Scanning charge-to-alanine mutagenesis has shown that K173, R174 and K177 in the C-terminal domain of TCS are involved in interacting with the P2, presumably through forming charge-charge interactions to the conserved DDD motif at the C-terminal tail of P2. A triple-alanine variant K173A/R174A/K177A of TCS, which fails to bind P2 and ribosomal stalk in vitro, was found to be 18-fold less active in inhibiting translation in rabbit reticulocyte lysate, suggesting that interaction with P-proteins is required for full activity of TCS. In an analogy to the role of stalk proteins in binding elongation factors, we propose that interaction with acidic ribosomal stalk proteins help TCS to locate its RNA substrate.
Figures

References
-
- Shaw PC, Lee KM, Wong KB. Recent advances in trichosanthin, a ribosome-inactivating protein with multiple pharmacological properties. Toxicon. 2005;45:683–689. - PubMed
-
- Endo Y, Tsurugi K. RNA N-glycosidase activity of ricin A-chain. Mechanism of action of the toxic lectin ricin on eukaryotic ribosomes. J. Biol. Chem. 1987;262:8128–8130. - PubMed
-
- Endo Y, Mitsui K, Motizuki M, Tsurugi K. The mechanism of action of ricin and related toxic lectins on eukaryotic ribosomes. The site and the characteristics of the modification in 28 S ribosomal RNA caused by the toxins. J. Biol. Chem. 1987;262:5908–5912. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
