

Exocytosis of ATP from astrocyte progenitors modulates spontaneous Ca2+ oscillations and cell migration
- PMID: 17309060
- PMCID: PMC2617704
- DOI: 10.1002/glia.20494
Exocytosis of ATP from astrocyte progenitors modulates spontaneous Ca2+ oscillations and cell migration
Abstract
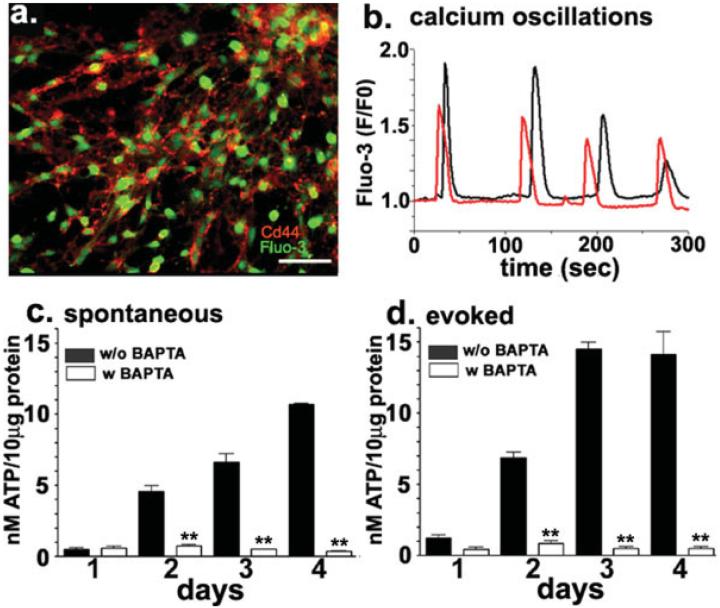
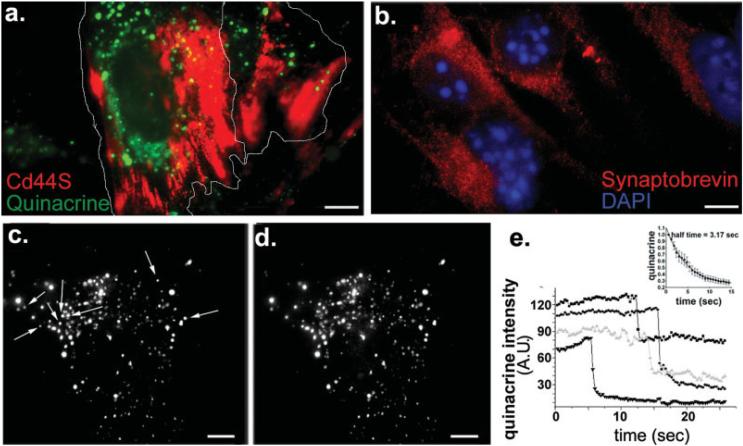
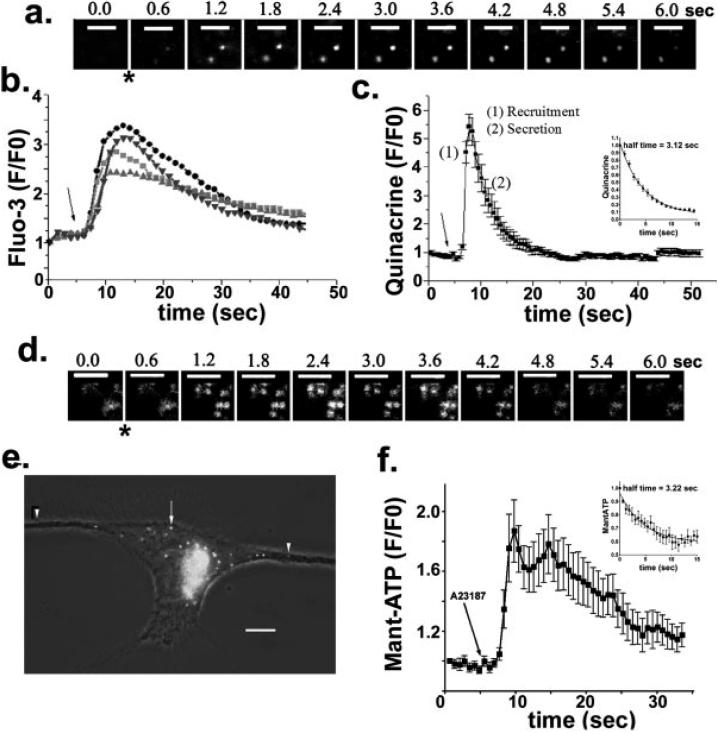
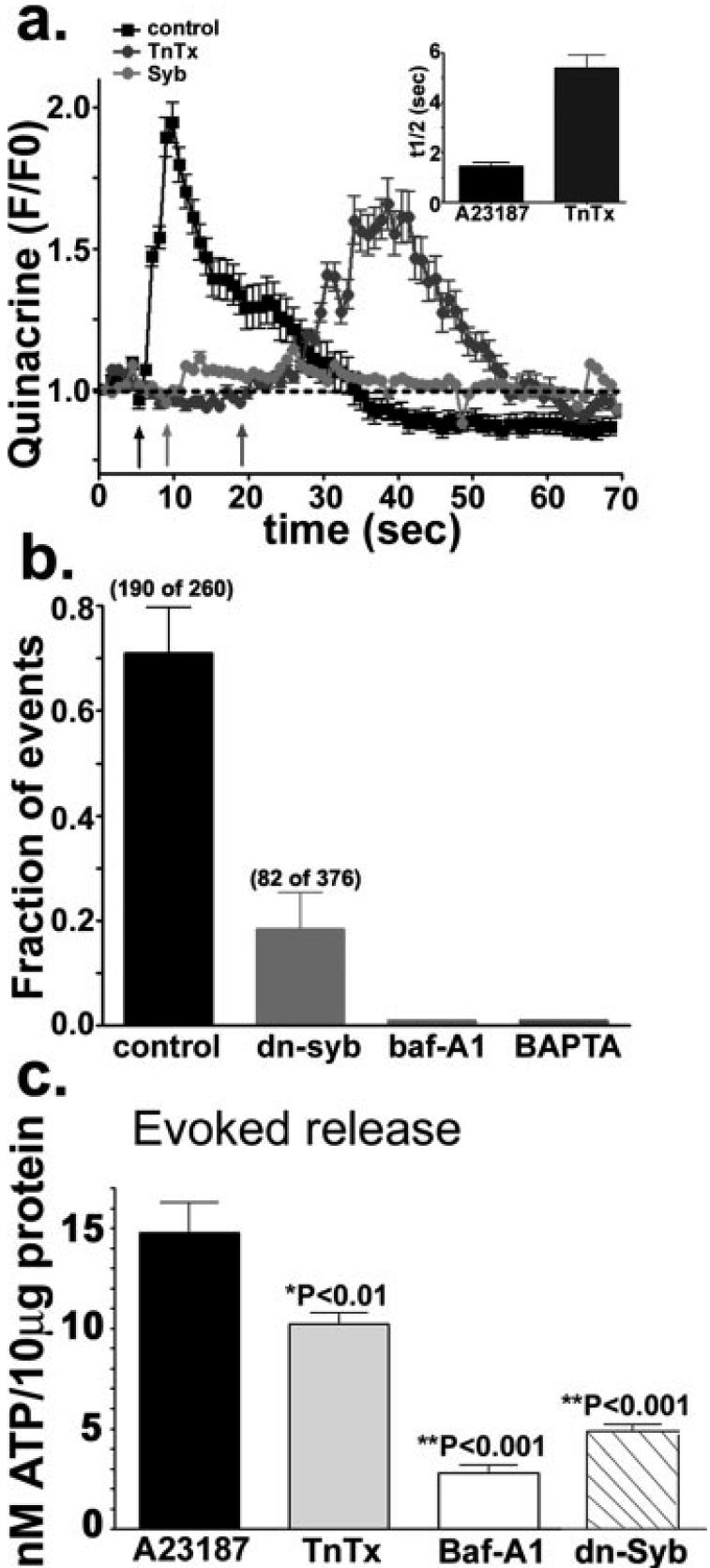
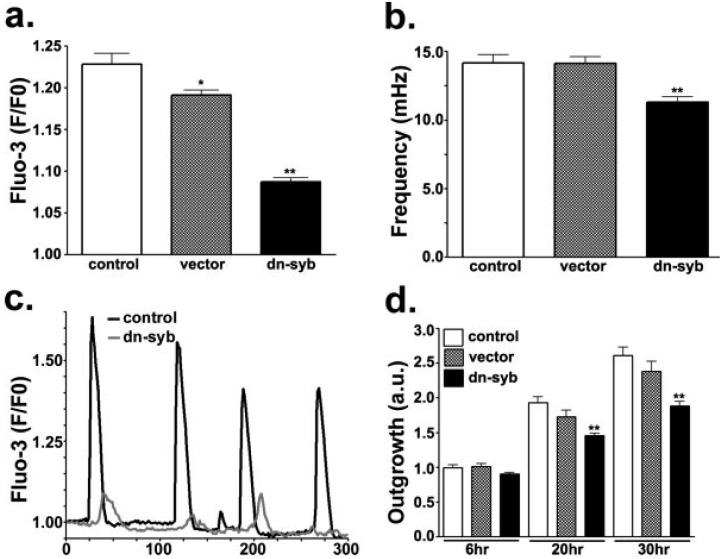
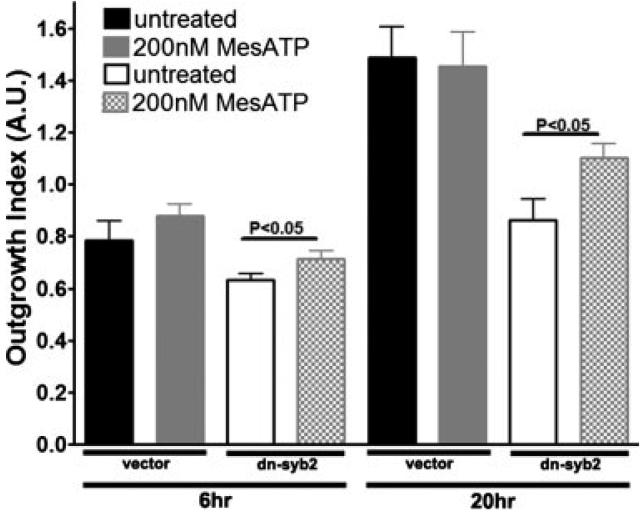
In the mature central nervous system (CNS) regulated secretion of ATP from astrocytes is thought to play a significant role in cell signaling. Whether such a mechanism is also operative in the developing nervous system and, if so, during which stage of development, has not been investigated. We have tackled this question using cells derived from reconstituted neurospheres, as well as brain explants of embryonic mice. Here, we show that in both models of neural cell development, astrocyte progenitors are competent for the regulated secretion of ATP-containing vesicles. We further document that this secretion is dependent on cytosolic Ca(2+) and the v-SNARE system, and takes place by exocytosis. Interference with ATP secretion alters spontaneous Ca(2+) oscillations and migration of neural progenitors. These data indicate that astrocyte progenitors acquire early in development the competence for regulated secretion of ATP, and that this event is implicated in the regulation of at least two cell functions, which are critical for the proper morphogenesis and functional maturation of the CNS.
Figures
Similar articles
-
Regulation of exocytotic protein expression and Ca2+-dependent peptide secretion in astrocytes.J Neurochem. 2009 Jul;110(1):143-56. doi: 10.1111/j.1471-4159.2009.06116.x. Epub 2009 May 15. J Neurochem. 2009. PMID: 19594665
-
Reduced expression of P2Y1 receptors in connexin43-null mice alters calcium signaling and migration of neural progenitor cells.J Neurosci. 2003 Dec 10;23(36):11444-52. doi: 10.1523/JNEUROSCI.23-36-11444.2003. J Neurosci. 2003. PMID: 14673009 Free PMC article.
-
Exocytosis of ATP from astrocytes modulates phasic and tonic inhibition in the neocortex.PLoS Biol. 2014 Jan;12(1):e1001747. doi: 10.1371/journal.pbio.1001747. Epub 2014 Jan 7. PLoS Biol. 2014. PMID: 24409095 Free PMC article.
-
Loose excitation-secretion coupling in astrocytes.Glia. 2016 May;64(5):655-67. doi: 10.1002/glia.22920. Epub 2015 Sep 11. Glia. 2016. PMID: 26358496 Free PMC article. Review.
-
Exocytosis in Astrocytes.Biomolecules. 2021 Sep 16;11(9):1367. doi: 10.3390/biom11091367. Biomolecules. 2021. PMID: 34572580 Free PMC article. Review.
Cited by
-
Neuroblast migration and P2Y(1) receptor mediated calcium signalling depend on 9-O-acetyl GD3 ganglioside.ASN Neuro. 2012 Sep 11;4(6):357-69. doi: 10.1042/AN20120035. ASN Neuro. 2012. PMID: 22894715 Free PMC article.
-
Astrocyte and Neuronal Pannexin1 Contribute Distinctly to Seizures.ASN Neuro. 2019 Jan-Dec;11:1759091419833502. doi: 10.1177/1759091419833502. ASN Neuro. 2019. PMID: 30862176 Free PMC article.
-
NTPDase2 and purinergic signaling control progenitor cell proliferation in neurogenic niches of the adult mouse brain.Stem Cells. 2015 Jan;33(1):253-64. doi: 10.1002/stem.1846. Stem Cells. 2015. PMID: 25205248 Free PMC article.
-
Transcriptome and metabolome analysis of crGART, a novel cell model of de novo purine synthesis deficiency: Alterations in CD36 expression and activity.PLoS One. 2021 Jul 20;16(7):e0247227. doi: 10.1371/journal.pone.0247227. eCollection 2021. PLoS One. 2021. PMID: 34283828 Free PMC article.
-
Point mutation in the mouse P2X7 receptor affects intercellular calcium waves in astrocytes.ASN Neuro. 2009 Apr 14;1(1):e00005. doi: 10.1042/AN20090001. ASN Neuro. 2009. PMID: 19570022 Free PMC article.
References
-
- Abbracchio MP. ATP and brain function. In: Jacobson KA, Jarvis MF, editors. Purinergic approaches in experimental therapeutics. Willey-Liss; New York: 1997. pp. 383–404.
-
- Abdipranoto A, Liu GJ, Werry EL, Bennett MR. Mechanisms of secretion of ATP from cortical astrocytes triggered by uridine triphosphate. Neuroreport. 2003;14:2177–2181. - PubMed
-
- Agresti C, Meomartini ME, Amadio S, Ambrosini E, Serafini B, Franchini L, Volonte C, Aloisi F, Visentin S. Metabotropic P2 receptor activation regulates oligodendrocyte progenitor migration and development. Glia. 2005;50:132–144. - PubMed
-
- Ahnert-Hilger G, Bigalke H. Molecular aspects of tetanus and botulin neurotoxin poisoning. Prog Neurobiol. 1995;46:83–96. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
