

Epigenetic activation of a subset of mRNAs by eIF4E explains its effects on cell proliferation
- PMID: 17311107
- PMCID: PMC1797416
- DOI: 10.1371/journal.pone.0000242
Epigenetic activation of a subset of mRNAs by eIF4E explains its effects on cell proliferation
Abstract
Background: Translation deregulation is an important mechanism that causes aberrant cell growth, proliferation and survival. eIF4E, the mRNA 5' cap-binding protein, plays a major role in translational control. To understand how eIF4E affects cell proliferation and survival, we studied mRNA targets that are translationally responsive to eIF4E.
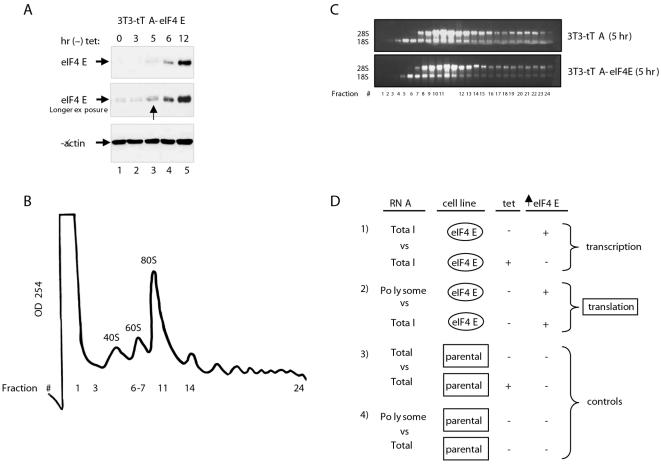
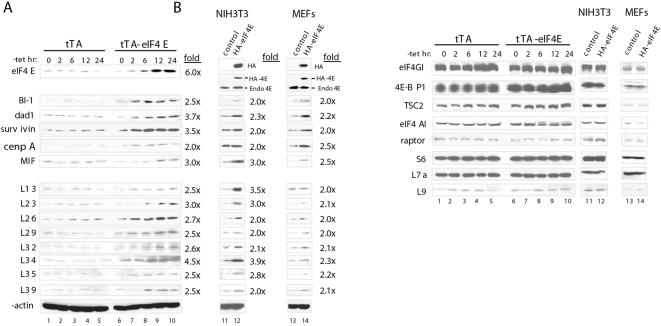
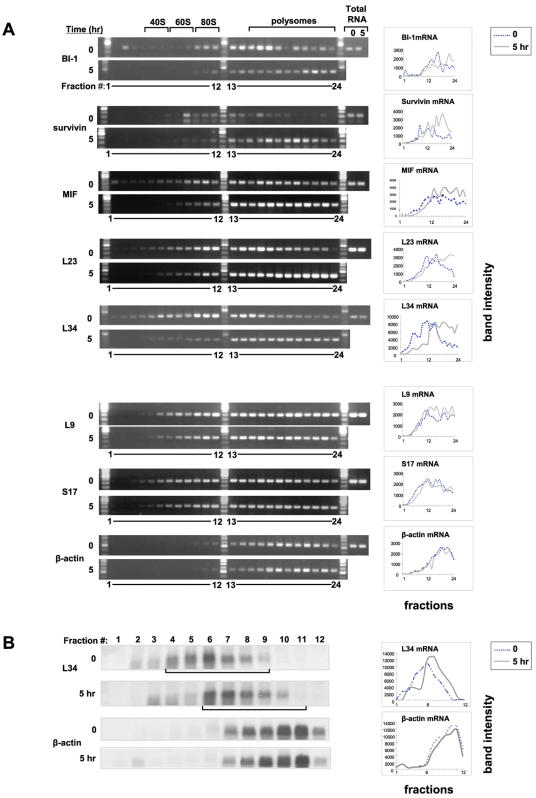
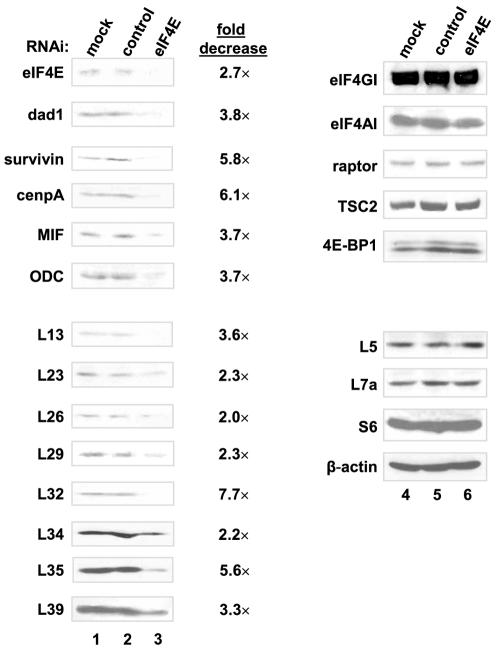
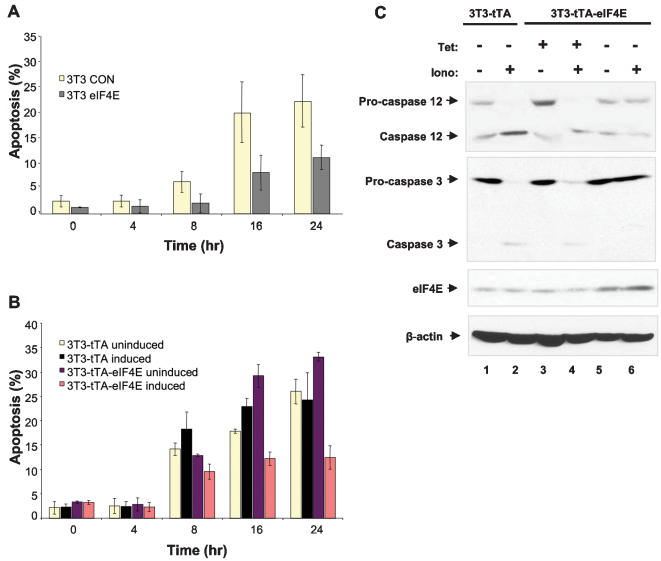
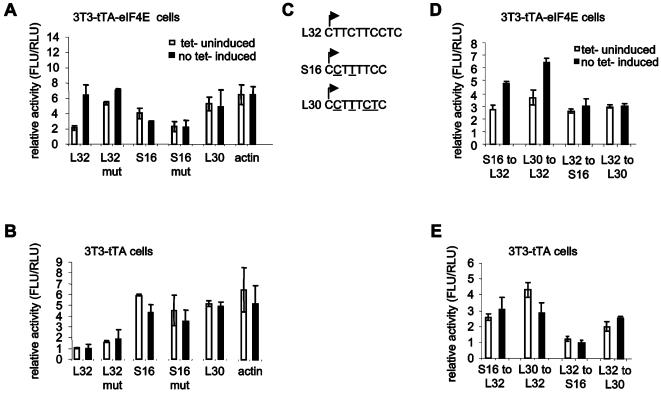
Methodology/principal findings: Microarray analysis of polysomal mRNA from an eIF4E-inducible NIH 3T3 cell line was performed. Inducible expression of eIF4E resulted in increased translation of defined sets of mRNAs. Many of the mRNAs are novel targets, including those that encode large- and small-subunit ribosomal proteins and cell growth-related factors. In addition, there was augmented translation of mRNAs encoding anti-apoptotic proteins, which conferred resistance to endoplasmic reticulum-mediated apoptosis.
Conclusions/significance: Our results shed new light on the mechanisms by which eIF4E prevents apoptosis and transforms cells. Downregulation of eIF4E and its downstream targets is a potential therapeutic option for the development of novel anti-cancer drugs.
Conflict of interest statement
Figures
References
-
- Thompson B, Wickens M, Kimble J. Translational Control in Development. In: Mathews M, Sonenberg N, Hershey J, editors. Translational Control in Biology and Medicine. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press; 2007.
-
- Schneider R, Sonenberg N. Translational Control in Cancer Development and Progression. In: Mathews M, Sonenberg N, Hershey J, editors. Translational Control in Biology and Medicine. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press; 2007.
-
- Tee AR, Blenis J. mTOR, translational control and human disease. Semin Cell Dev Biol. 2005;16:29–37. - PubMed
-
- Dua K, Williams TM, Beretta L. Translational control of the proteome: relevance to cancer. Proteomics. 2001;1:1191–1199. - PubMed
-
- Holcik M, Sonenberg N. Translational control in stress and apoptosis. Nat Rev Mol Cell Biol. 2005;6:318–327. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
