

Influence of translation efficiency of homologous viral proteins on the endogenous presentation of CD8+ T cell epitopes
- PMID: 17312009
- PMCID: PMC2137904
- DOI: 10.1084/jem.20062508
Influence of translation efficiency of homologous viral proteins on the endogenous presentation of CD8+ T cell epitopes
Abstract
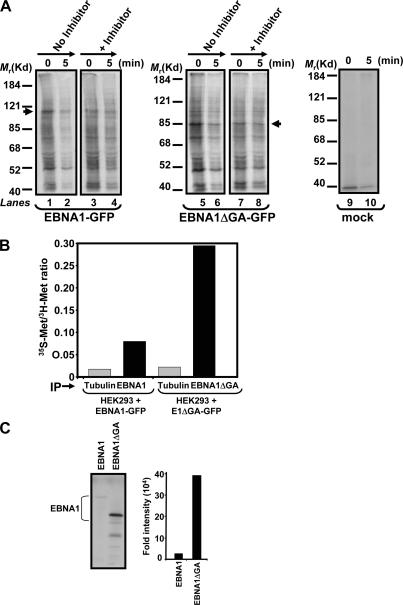
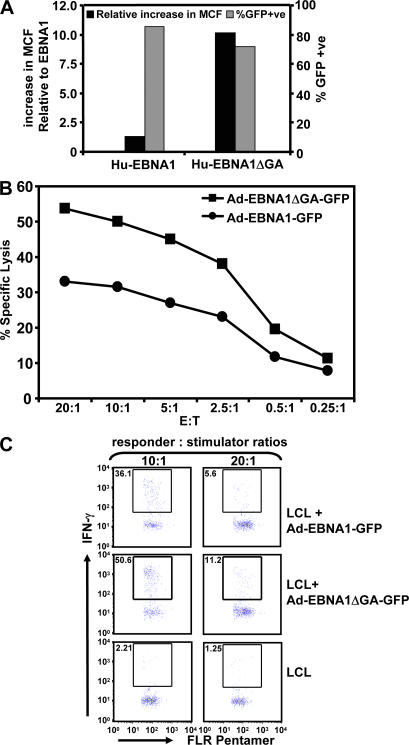
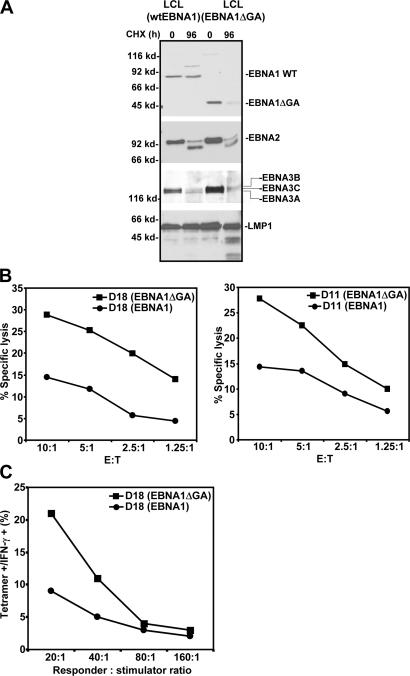
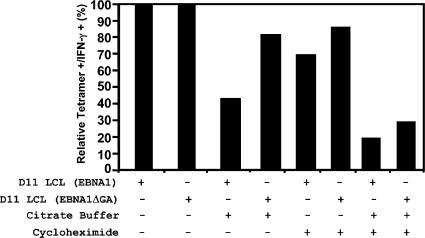
A significant proportion of endogenously processed CD8(+) T cell epitopes are derived from newly synthesized proteins and rapidly degrading polypeptides (RDPs). It has been hypothesized that the generation of rapidly degrading polypeptides and CD8(+) T cell epitopes from these RDP precursors may be influenced by the efficiency of protein translation. Here we address this hypothesis by using the Epstein-Barr virus-encoded nuclear antigen 1 protein (EBNA1), with or without its internal glycine-alanine repeat sequence (EBNA1 and EBNA1DeltaGA, respectively), which display distinct differences in translation efficiency. We demonstrate that RDPs constitute a significant proportion of newly synthesized EBNA1 and EBNA1DeltaGA and that the levels of RDPs produced by each of these proteins directly correlate with the translation efficiency of either EBNA1 or EBNA1DeltaGA. As a consequence, a higher number of major histocompatibility complex-peptide complexes can be detected on the surface of cells expressing EBNA1DeltaGA, and these cells are more efficiently recognized by virus-specific cytotoxic T lymphocytes compared to the full-length EBNA1. More importantly, we also demonstrate that the endogenous processing of these CD8(+) T cell epitopes is predominantly determined by the rate at which the RDPs are generated rather than the intracellular turnover of these proteins.
Figures
Similar articles
-
Translation efficiency of EBNA1 encoded by lymphocryptoviruses influences endogenous presentation of CD8+ T cell epitopes.Eur J Immunol. 2007 Feb;37(2):328-37. doi: 10.1002/eji.200636153. Eur J Immunol. 2007. PMID: 17236233
-
Endogenous presentation of CD8+ T cell epitopes from Epstein-Barr virus-encoded nuclear antigen 1.J Exp Med. 2004 May 17;199(10):1421-31. doi: 10.1084/jem.20040191. J Exp Med. 2004. PMID: 15148340 Free PMC article.
-
Inhibition of antigen presentation by the glycine/alanine repeat domain is not conserved in simian homologues of Epstein-Barr virus nuclear antigen 1.J Virol. 1999 Sep;73(9):7381-9. doi: 10.1128/JVI.73.9.7381-7389.1999. J Virol. 1999. PMID: 10438828 Free PMC article.
-
Epstein-Barr virus evasion of CD8(+) and CD4(+) T cell immunity via concerted actions of multiple gene products.Semin Cancer Biol. 2008 Dec;18(6):397-408. doi: 10.1016/j.semcancer.2008.10.008. Epub 2008 Oct 25. Semin Cancer Biol. 2008. PMID: 18977445 Review.
-
Avoiding proteasomal processing: the case of EBNA1.Curr Top Microbiol Immunol. 2002;269:23-36. doi: 10.1007/978-3-642-59421-2_2. Curr Top Microbiol Immunol. 2002. PMID: 12224511 Review.
Cited by
-
Messenger RNA sequence rather than protein sequence determines the level of self-synthesis and antigen presentation of the EBV-encoded antigen, EBNA1.PLoS Pathog. 2012 Dec;8(12):e1003112. doi: 10.1371/journal.ppat.1003112. Epub 2012 Dec 27. PLoS Pathog. 2012. PMID: 23300450 Free PMC article.
-
Modified Vaccinia Virus Ankara Can Induce Optimal CD8+ T Cell Responses to Directly Primed Antigens Depending on Vaccine Design.J Virol. 2019 Oct 15;93(21):e01154-19. doi: 10.1128/JVI.01154-19. Print 2019 Nov 1. J Virol. 2019. PMID: 31375596 Free PMC article.
-
Re-examining class-I presentation and the DRiP hypothesis.Trends Immunol. 2014 Apr;35(4):144-52. doi: 10.1016/j.it.2014.01.002. Epub 2014 Feb 21. Trends Immunol. 2014. PMID: 24566257 Free PMC article. Review.
-
Decreased EBNA-1-specific CD8+ T cells in patients with Epstein-Barr virus-associated nasopharyngeal carcinoma.Proc Natl Acad Sci U S A. 2009 Mar 3;106(9):3318-23. doi: 10.1073/pnas.0813320106. Epub 2009 Feb 11. Proc Natl Acad Sci U S A. 2009. PMID: 19211798 Free PMC article.
-
Autorepression of Epstein-Barr virus nuclear antigen 1 expression by inhibition of pre-mRNA processing.J Virol. 2008 Feb;82(4):1679-87. doi: 10.1128/JVI.02142-07. Epub 2007 Dec 12. J Virol. 2008. PMID: 18077719 Free PMC article.
References
-
- Yewdell, J.W., and S.M. Haeryfar. 2005. Understanding presentation of viral antigens to CD8+ T cells in vivo: the key to rational vaccine design. Annu. Rev. Immunol. 23:651–682. - PubMed
-
- Yewdell, J.W., E. Reits, and J. Neefjes. 2003. Making sense of mass destruction: quantitating MHC class I antigen presentation. Nat. Rev. Immunol. 3:952–961. - PubMed
-
- Yewdell, J.W., U. Schubert, and J.R. Bennink. 2001. At the crossroads of cell biology and immunology: DRiPs and other sources of peptide ligands for MHC class I molecules. J. Cell Sci. 114:845–851. - PubMed
-
- Goldberg, A.L., P. Cascio, T. Saric, and K.L. Rock. 2002. The importance of the proteasome and subsequent proteolytic steps in the generation of antigenic peptides. Mol. Immunol. 39:147–164. - PubMed
-
- Goldberg, A.L., and K.L. Rock. 1992. Proteolysis, proteasomes and antigen presentation. Nature. 357:375–379. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
