

Identification of a Fab interaction footprint site on an icosahedral virus by cryoelectron microscopy and X-ray crystallography
- PMID: 1731227
- PMCID: PMC4152847
- DOI: 10.1038/355275a0
Identification of a Fab interaction footprint site on an icosahedral virus by cryoelectron microscopy and X-ray crystallography
Abstract
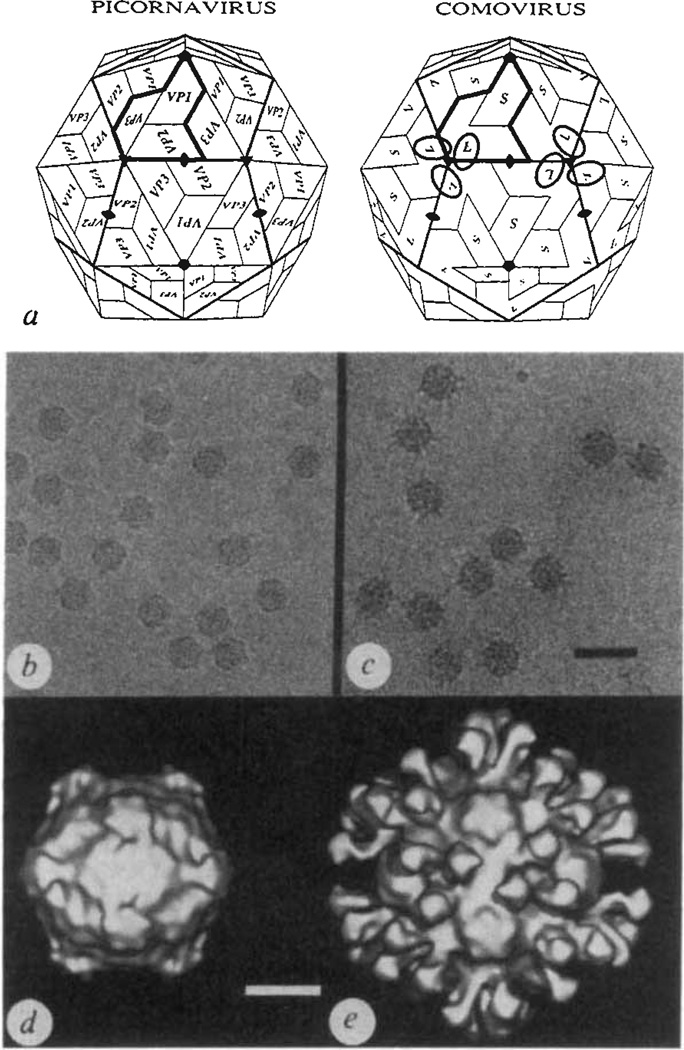
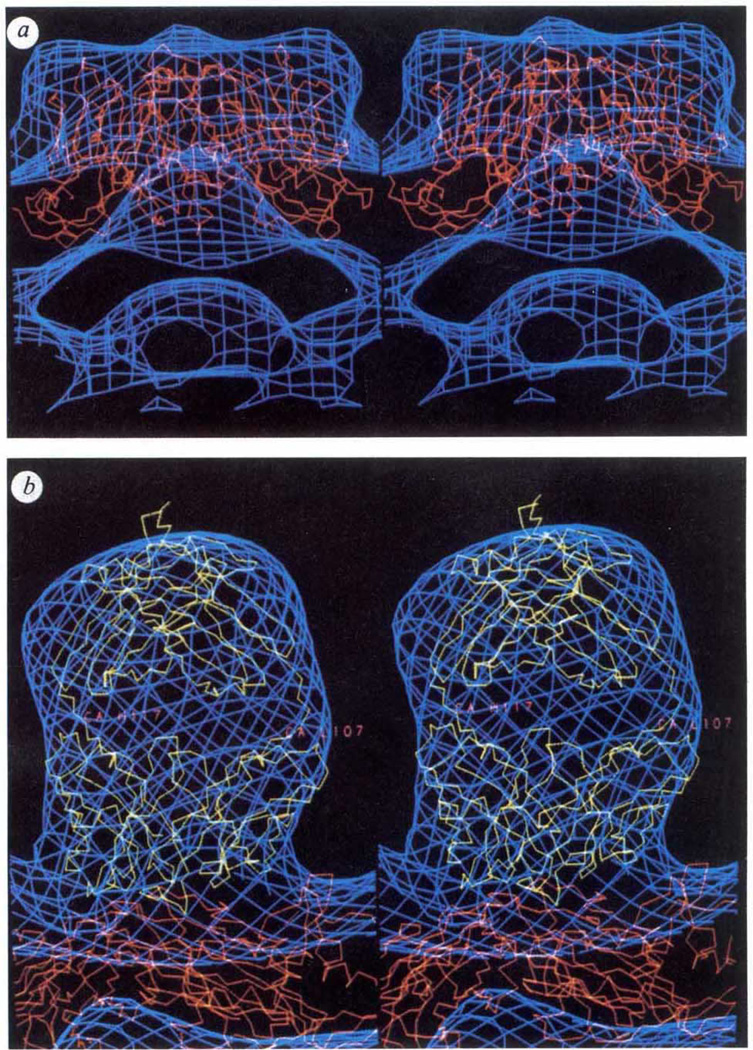
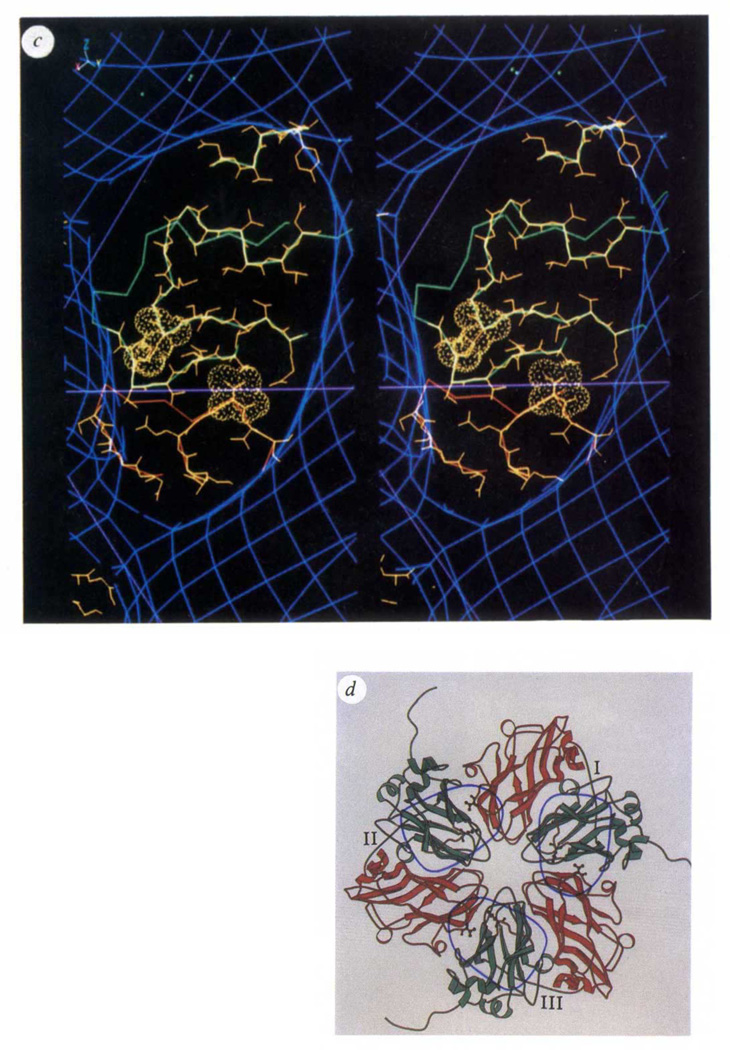
Biological processes frequently require the formation of multi-protein or nucleoprotein complexes. Some of these complexes have been produced in homogeneous form, crystallized, and analysed at high resolution by X-ray crystallography (for example, see refs 1-3). Most, however, are too large or too unstable to crystallize. Individual components of such complexes can often be purified and analysed by crystallography. Here we report how the coordinated application of cryoelectron microscopy, three-dimensional image reconstruction, and X-ray crystallography provides a powerful approach to study large, unstable macromolecular complexes. Three-dimensional reconstructions of native cowpea mosaic virus (CMPV) and a complex of CPMV saturated with a Fab fragment of a monoclonal antibody against the virus have been determined at 23 A resolution from low-irradiation images of unstained, frozen-hydrated samples. Despite the nominal resolution of the complex, the physical footprint of the Fab on the capsid surface and the orientation and position of the Fab have been determined to within a few ångstroms by fitting atomic models of CPMV4 and Fab (Kol)5 to reconstructed density maps.
Figures
References
-
- Davies DR, Padlan EA, Sheriff S. A. Rev. Biochem. 1990;59:439–473. - PubMed
-
- Ruff M, et al. Science. 1991;252:1682–1689. - PubMed
-
- Rossmann MG, Johnson JE. A. Rev. Biochem. 1989;58:533–573. - PubMed
-
- Chen Z, Stauffacher CV, Johnson JE. Semin. Virol. 1990;1:453–466.
-
- Marquart M, Deisenhofer J, Huber R, Palm W. J. molec. Biol. 1980;141:369–391. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
