

Sparse and selective odor coding by mitral/tufted neurons in the main olfactory bulb
- PMID: 17314304
- PMCID: PMC6673545
- DOI: 10.1523/JNEUROSCI.3779-06.2007
Sparse and selective odor coding by mitral/tufted neurons in the main olfactory bulb
Abstract
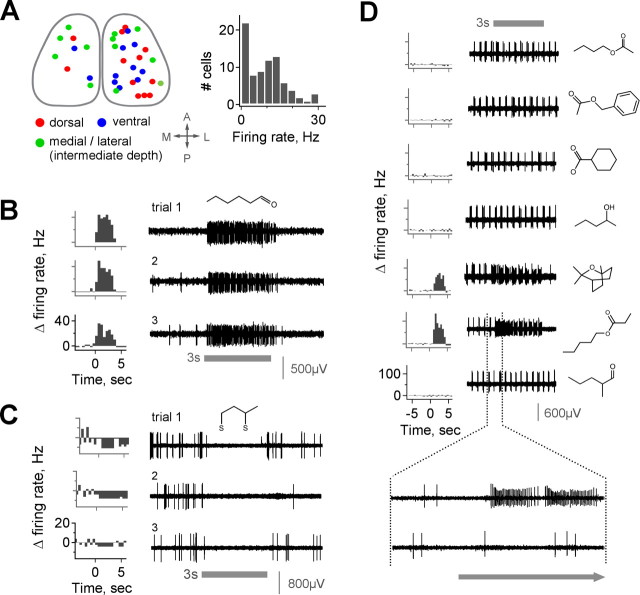
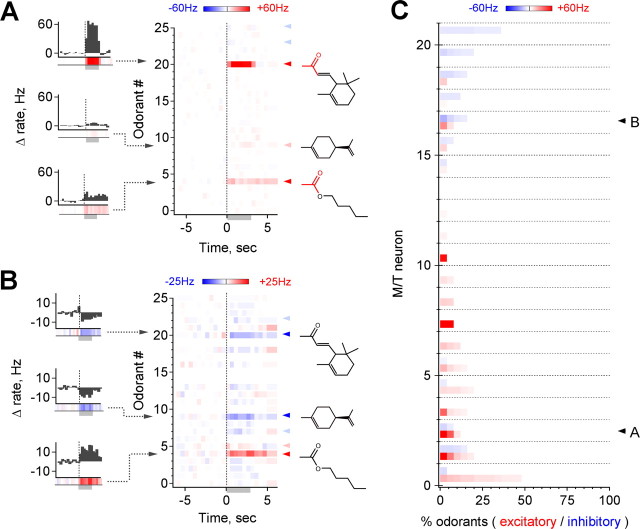
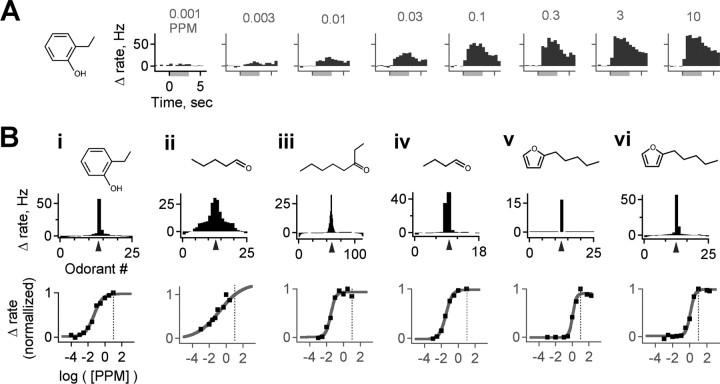
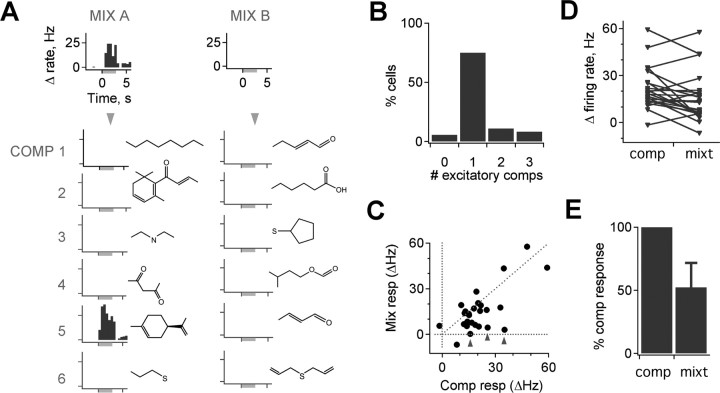
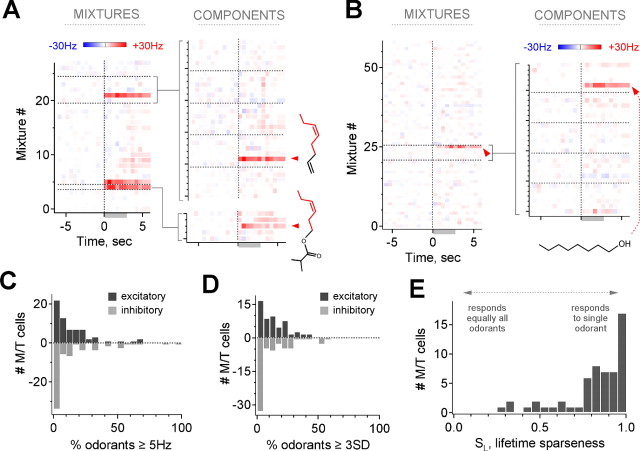
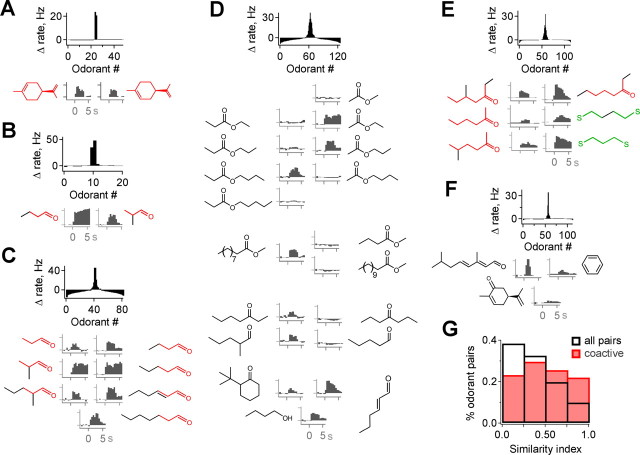
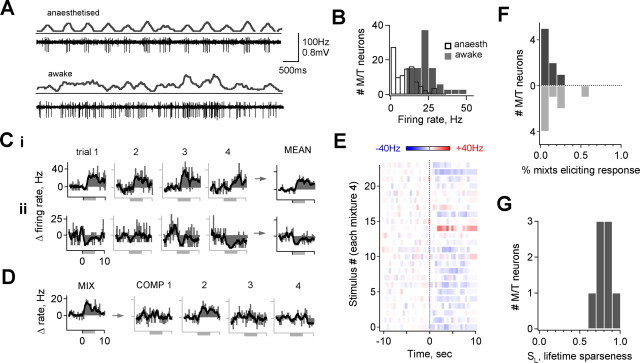
The mammalian olfactory system recognizes an enormous variety of odorants carrying a wide range of important behavioral cues. In the main olfactory bulb (MOB), odorants are ultimately represented through the action potential activity of mitral/tufted cells (M/Ts), whose selectivity and tuning to odorant molecules are therefore fundamental determinants of MOB sensory coding. However, the sheer number and diversity of discrete olfactory stimuli has been a major barrier to comprehensively evaluating M/T selectivity. To address this issue, we assessed M/T odorant responses in anesthetized mice to a 348-odorant panel widely and systematically distributed throughout chemical space, presented both individually and in mixtures at behaviorally relevant concentrations. We found that M/T activation by odorants was markedly selective, with neurons responding robustly, sensitively, and reliably to only a highly restricted subset of stimuli. Multiple odorants activating a single neuron commonly shared clear structural similarity, but M/T tuning also frequently extended beyond obviously defined chemical categories. Cells typically responded to effective compounds presented both individually and in mixtures, although firing rates evoked by mixtures typically showed partial suppression. Response selectivity was further confirmed in awake animals by chronic recordings of M/Ts. These data indicate that individual M/Ts encode specific odorant attributes shared by only a small fraction of compounds and imply that the MOB relays the collective molecular features of an odorant stimulus through a restricted set of M/Ts, each narrowly tuned to a particular stimulus characteristic.
Figures
References
-
- Aungst JL, Heyward PM, Puche AC, Karnup SV, Hayar A, Szabo G, Shipley MT. Centre-surround inhibition among olfactory bulb glomeruli. Nature. 2003;426:623–629. - PubMed
-
- Bargmann C. Comparative chemosensation from receptors to ecology. Nature. 2006;444:295–301. - PubMed
-
- Bhalla US, Bower JM. Multiday recordings from olfactory bulb neurons in awake freely moving rats: spatially and temporally organized variability in odorant response properties. J Comput Neurosci. 1997;4:221–256. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources