

Fatty acid remodeling of GPI-anchored proteins is required for their raft association
- PMID: 17314402
- PMCID: PMC1838968
- DOI: 10.1091/mbc.e06-10-0885
Fatty acid remodeling of GPI-anchored proteins is required for their raft association
Abstract
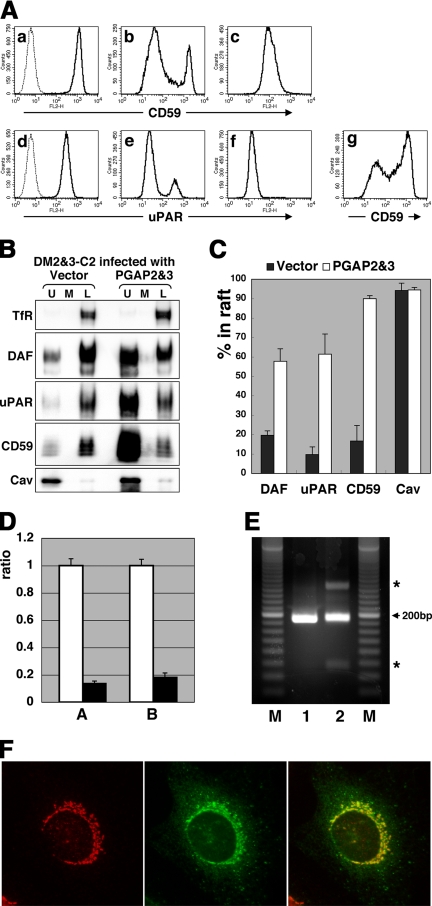
Whereas most of the cellular phosphatidylinositol (PI) contain unsaturated fatty chains and are excluded from rafts, GPI-anchored proteins (APs) unusually contain two saturated fatty chains in their PI moiety, and they are typically found within lipid rafts. However, the origin of the saturated chains and whether they are essential for raft association are unclear. Here, we report that GPI-APs, with two saturated fatty chains, are generated from those bearing an unsaturated chain by fatty acid remodeling that occurs most likely in the Golgi and requires post-GPI-attachment to proteins (PGAP)2 and PGAP3. The surface GPI-APs isolated from the PGAP2 and -3 double-mutant Chinese hamster ovary (CHO) cells had unsaturated chains, such as oleic, arachidonic, and docosatetraenoic acids in the sn-2 position, whereas those from wild-type CHO cells had exclusively stearic acid, a saturated chain, indicating that the sn-2 chain is exchanged to a saturated chain. We then assessed the association of GPI-APs with lipid rafts. Recovery of unremodeled GPI-APs from the double-mutant cells in the detergent-resistant membrane fraction was very low, indicating that GPI-APs become competent to be incorporated into lipid rafts by PGAP3- and PGAP2-mediated fatty acid remodeling. We also show that the remodeling requires the preceding PGAP1-mediated deacylation from inositol of GPI-APs in the endoplasmic reticulum.
Figures




References
-
- Benting J., Rietveld A., Ansorge I., Simons K. Acyl and alkyl chain length of GPI-anchors is critical for raft association in vitro. FEBS Lett. 1999;462:47–50. - PubMed
-
- Brewis I. A., Ferguson M. A., Mehlert A., Turner A. J., Hooper N. M. Structures of the glycosyl-phosphatidylinositol anchors of porcine and human renal membrane dipeptidase. Comprehensive structural studies on the porcine anchor and interspecies comparison of the glycan core structures. J. Biol. Chem. 1995;270:22946–22956. - PubMed
-
- Brown D. A., Rose J. K. Sorting of GPI-anchored proteins to glycolipid-enriched membrane subdomains during transport to the apical cell surface. Cell. 1992;68:533–544. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
