

Review
doi: 10.1038/nature05659.
Scratching the surface of skin development
Affiliations
- PMID: 17314969
- PMCID: PMC2405926
- DOI: 10.1038/nature05659
Item in Clipboard
Review
Scratching the surface of skin development
Nature.
.
Abstract
The epidermis and its appendages develop from a single layer of multipotent embryonic progenitor keratinocytes. Embryonic stem cells receive cues from their environment that instruct them to commit to a particular differentiation programme and generate a stratified epidermis, hair follicles or sebaceous glands. Exciting recent developments have focused on how adult skin epithelia maintain populations of stem cells for use in the natural cycles of hair follicle regeneration and for re-epithelialization in response to wounding.
Figures

a, In the absence of Wnt signalling, ectodermal progenitors respond to FGFs, downregulate BMP signalling and progress towards neurogenesis. Wnt signalling blocks the ability of early ectodermal progenitor cells to respond to FGFs, allowing them to respond to BMP signalling and adopt an epidermal fate. As development progresses, single-layered embryonic epidermis expresses Wnts. Some cells fail to respond to Wnts, and these become fated to become epidermal cells through BMP, EGF and Notch signalling. The cells that do respond to Wnt signalling also receive underlying FGF and BMP inhibitory signals from the mesenchyme and, together, these signals instruct the cells to make an appendage,-. Collectively, the inhibition of BMP inhibitory signals and Wnt activating signals produce the hair placode,,. Additional dermal messages from below further instruct the placodes to make the follicle.

a, Schematic of the hair bulb. Rapidly proliferating matrix cells at the base (blue) give rise to seven differentiation-specific lineages. The various genetic markers for these lineages are shown. Matrix cells express the proliferative marker Ki67. As these cells differentiate, they express keratins differentially. Ch, hair shaft cuticle; Co, cortex; Cp, companion layer; DP, dermal papilla; Me, medulla,. b, Ultrathin section of hair bulb. The single vertical layer of core cells corresponds to the dermal papilla. Note the onion-skin-like organization of matrix cells and their differentiating progeny at the base of the follicle. (Image courtesy of H. A. Pasolli, The Rockefeller University, New York.)

The stages of the hair cycle are depicted, starting from the first postnatal anagen, when the hair shaft is growing and protruding through the skin surface. Follicles progress synchronously to the destructive (catagen) phase, during which the lower two-thirds of the follicle undergo apoptosis and regress. The dermal papilla is brought to rest below the bulge-stem-cell compartment, and after the resting (telogen) phase, a critical threshold of activating factors is reached and the stem cells become activated to regrow the hair.

The bulge contains infrequently cycling, label-retaining cells, which include multipotent stem cells (green) that can generate the new hair follicle during cycling and repair the epidermis on injury. The bulge is in a specialized niche, surrounded by other cell types, which together provide cues that maintain these cells in an undifferentiated, quiescent state. For stem cells to be activated, the niche environment must change.
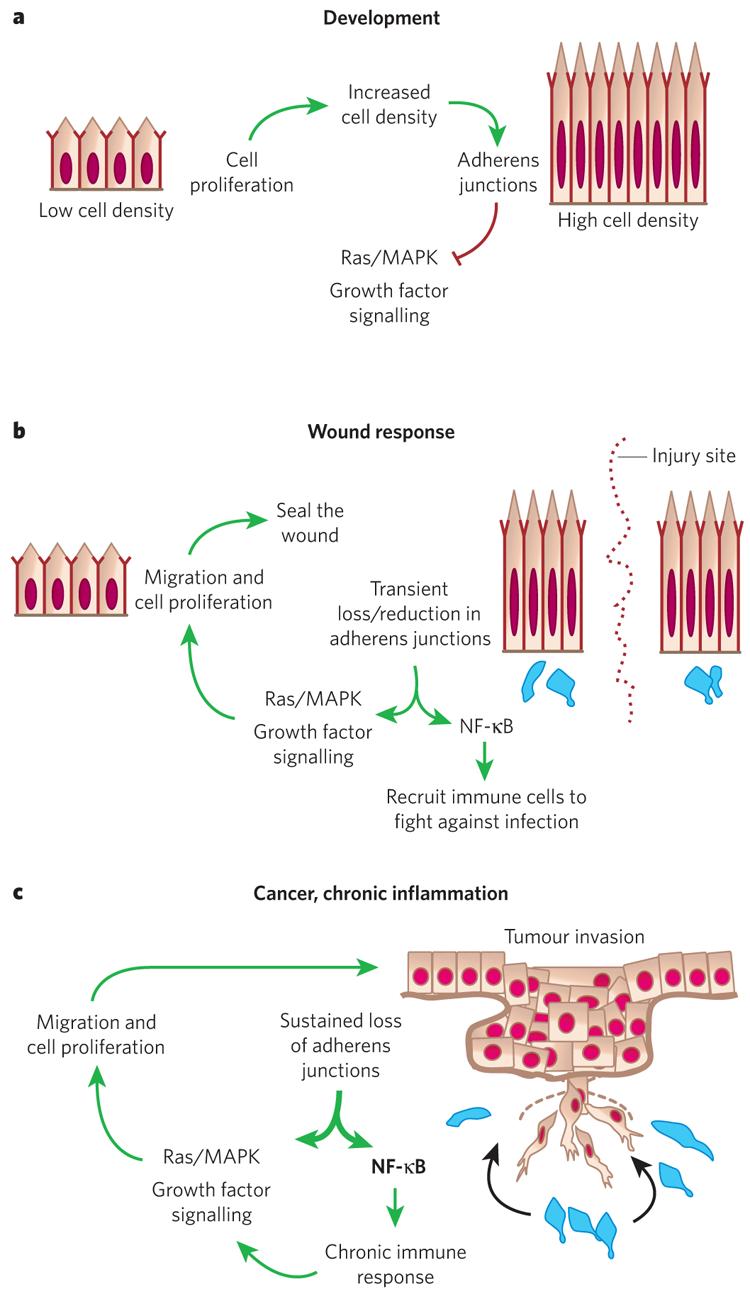
a, During development, basal epidermal cell density is low and cells proliferate. As cells increase in density the number of adherens junctions at cell–cell borders increases. This provides a regulatory feedback loop to control Ras–MAPK activity and cell proliferation. b, In a wound response, adherens junctions are severed at the wound site, stimulating the Ras–MAPK pathway and triggering a hyperproliferative response and cell migration. Adherens junction protein levels also affect NF-κB, which, when active, results in the recruitment of immune cells (blue) to protect against infection. After wound repair, regulatory feedback pathways cause these processes to be dampened and the system returns to normal. c, Cancer and chronic inflammatory disorders can lead to mutations and/or permanent changes in adherens junction proteins. This results in a break in the regulatory circuitry, and the proliferative and NF-κB responses become constitutively activated.

Symmetric and asymmetric cell division have been observed in the epidermis, and both are likely to contribute to epidermal homeostasis. Models of how they might do so are shown. Both are predicated on the basis of cellular attachment to an underlying basement membrane providing the main means by which basal epidermal progenitor cells are able to preserve their mitotic capability. a, Symmetric division is thought to supply new cells through stratification (the formation of layers) and differentiation by delamination (the detachment of a cell from the basement membrane). b, Asymmetric division is believed to contribute through stratification and differentiation by asymmetric partitioning of cellular contents into the two daughter cells.
References
-
- Stern CD. Neural induction: old problem, new findings, yet more questions. Development. 2005;132:2007–2021. - PubMed
-
- M'Boneko V, Merker HJ. Development and morphology of the periderm of mouse embryos (days 9–12 of gestation) Acta Anat. (Basel) 1988;133:325–336. - PubMed
-
- Atit R, et al. β-catenin activation is necessary and sufficient to specify the dorsal dermal fate in the mouse. Dev. Biol. 2006;296:164–176. - PubMed
-
- Hardy MH. The secret life of the hair follicle. Trends Genet. 1992;8:55–61. - PubMed
-
- Olivera-Martinez I, Thelu J, Dhouailly D. Molecular mechanisms controlling dorsal dermis generation from the somitic dermomyotome. Int. J. Dev. Biol. 2004;48:93–101. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
