

Trogocytosis-based generation of suppressive NK cells
- PMID: 17318190
- PMCID: PMC1817622
- DOI: 10.1038/sj.emboj.7601570
Trogocytosis-based generation of suppressive NK cells
Abstract
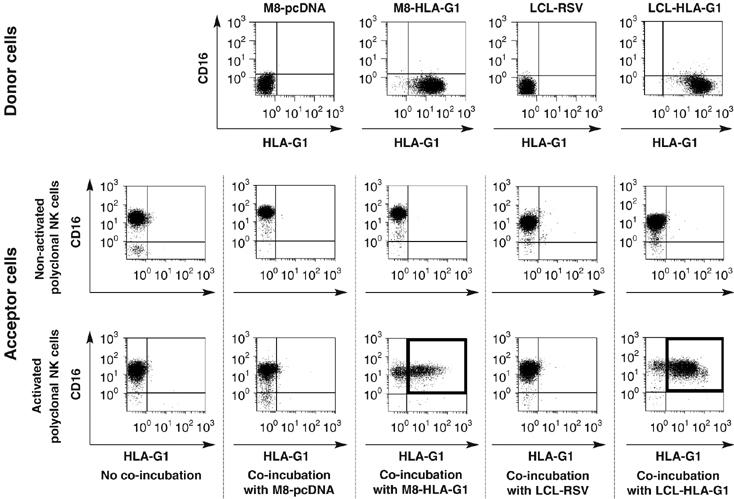
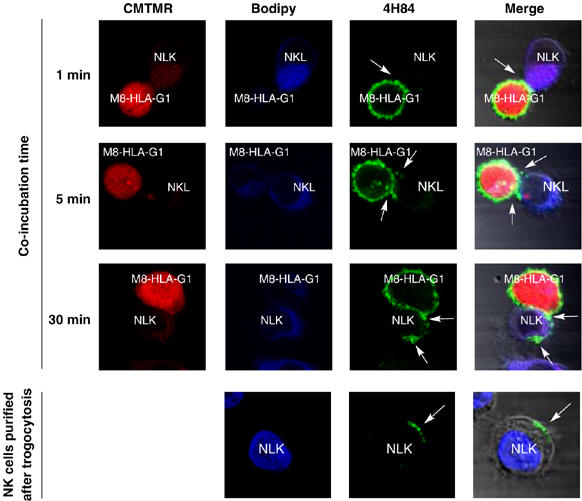
Trogocytosis is a fast uptake of membranes and associated molecules from one cell by another. Trogocytosis between natural killer (NK) cells and tumors is already described, but the functional relevance of NK-tumor targets material exchange is unclear. We investigated whether the immunosuppressive molecule HLA-G that is commonly expressed by tumors in vivo and known to block NK cytolytic function, could be transferred from tumor cells to NK cells, and if this transfer had functional consequences. We show that activated NK cells acquire HLA-G1 from tumor cells, and that upon this acquisition, NK cells stop proliferating, are no longer cytotoxic, and behave as suppressor cells. Such cells can inhibit other NK cells' cytotoxic function and protect NK-sensitive tumor cells from cytolysis. These data are the first demonstration that trogocytosis of HLA-G1 can be a major mechanism of immune escape that acts through effector cells made to act as suppressor cells locally, temporarily, but efficiently. The broader consequences of membrane sharing between immune and non-immune cells on the function of effectors and the outcome of immune responses are discussed.
Figures





References
-
- Bahri R, Hirsch F, Josse A, Rouas-Freiss N, Bidere N, Vasquez A, Carosella ED, Charpentier B, Durrbach A (2006) Soluble HLA-G inhibits cell cycle progression in human alloreactive T lymphocytes. J Immunol 176: 1331–1339 - PubMed
-
- Carosella ED, Moreau P, Le Maoult J, Le Discorde M, Dausset J, Rouas-Freiss N (2003) HLA-G molecules: from maternal–fetal tolerance to tissue acceptance. Adv Immunol 81: 199–252 - PubMed
-
- Game DS, Rogers NJ, Lechler RI (2005) Acquisition of HLA-DR and costimulatory molecules by T cells from allogeneic antigen presenting cells. Am J Transplant 7: 1614–1625 - PubMed
-
- Horuzsko A, Lenfant F, Munn DH, Mellor AL (2001) Maturation of antigen-presenting cells is compromised in HLA-G transgenic mice. Int Immunol 13: 385–394 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
