

Structure and interactions of the first three RNA recognition motifs of splicing factor prp24
- PMID: 17320109
- PMCID: PMC1939982
- DOI: 10.1016/j.jmb.2007.01.078
Structure and interactions of the first three RNA recognition motifs of splicing factor prp24
Abstract
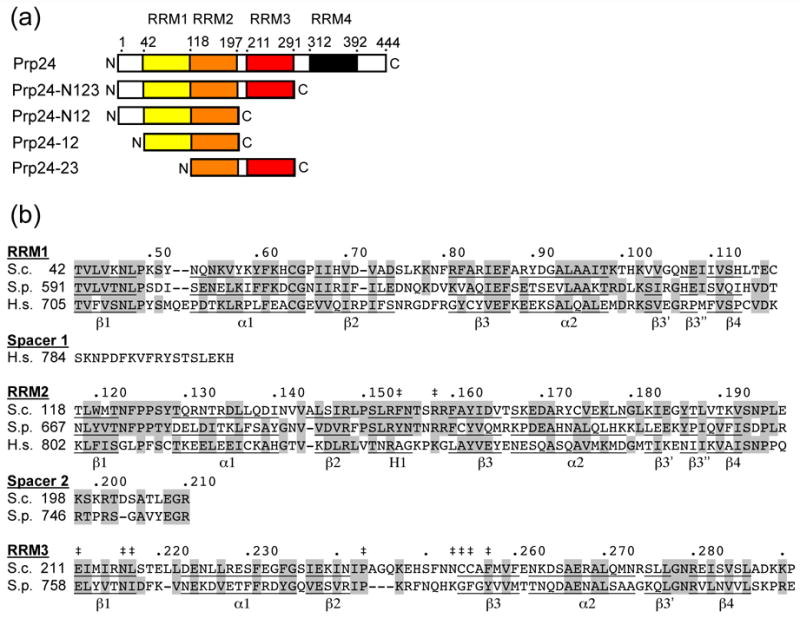
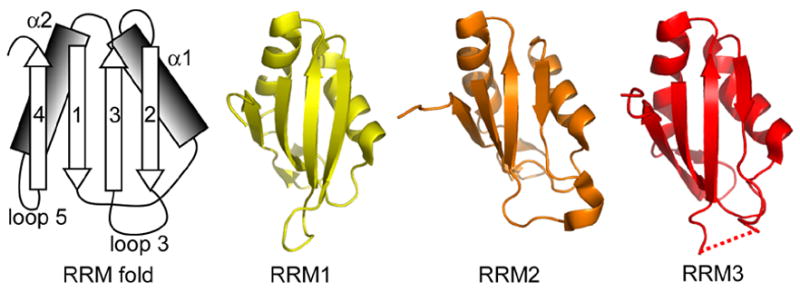
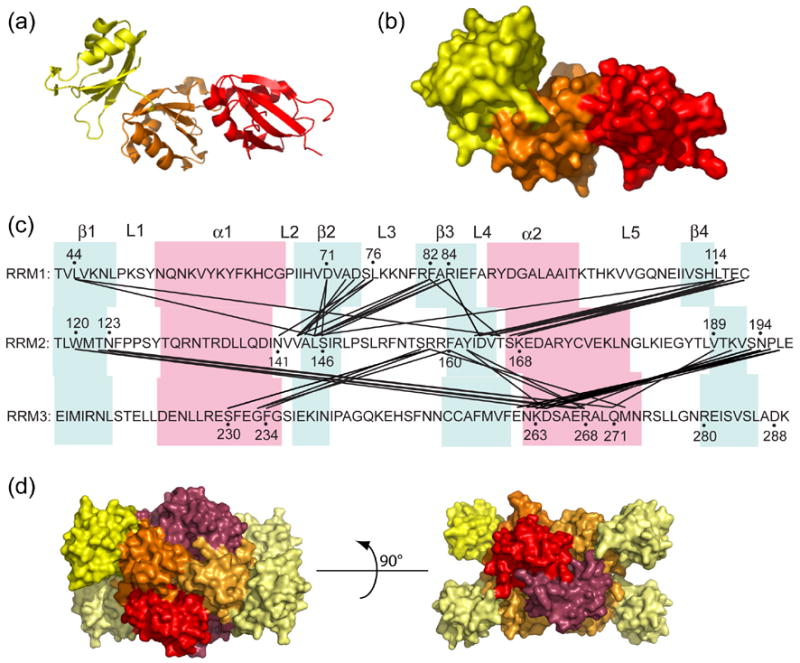
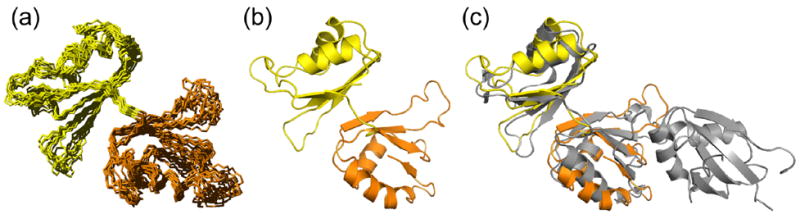
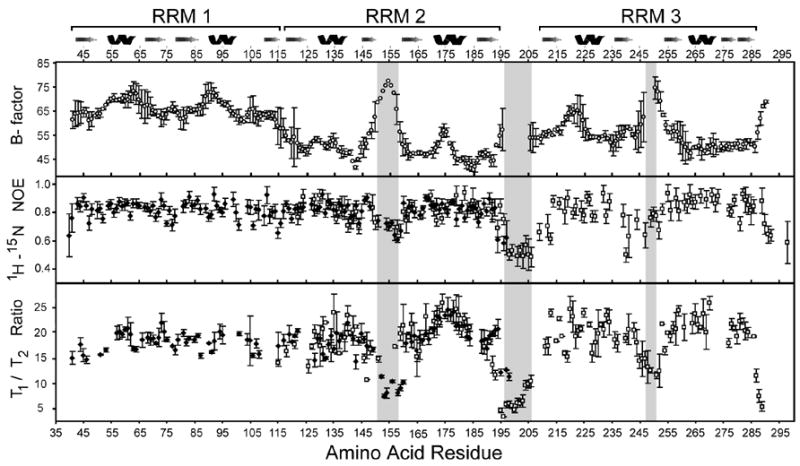
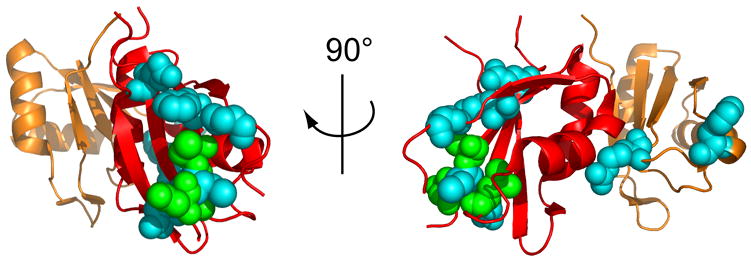
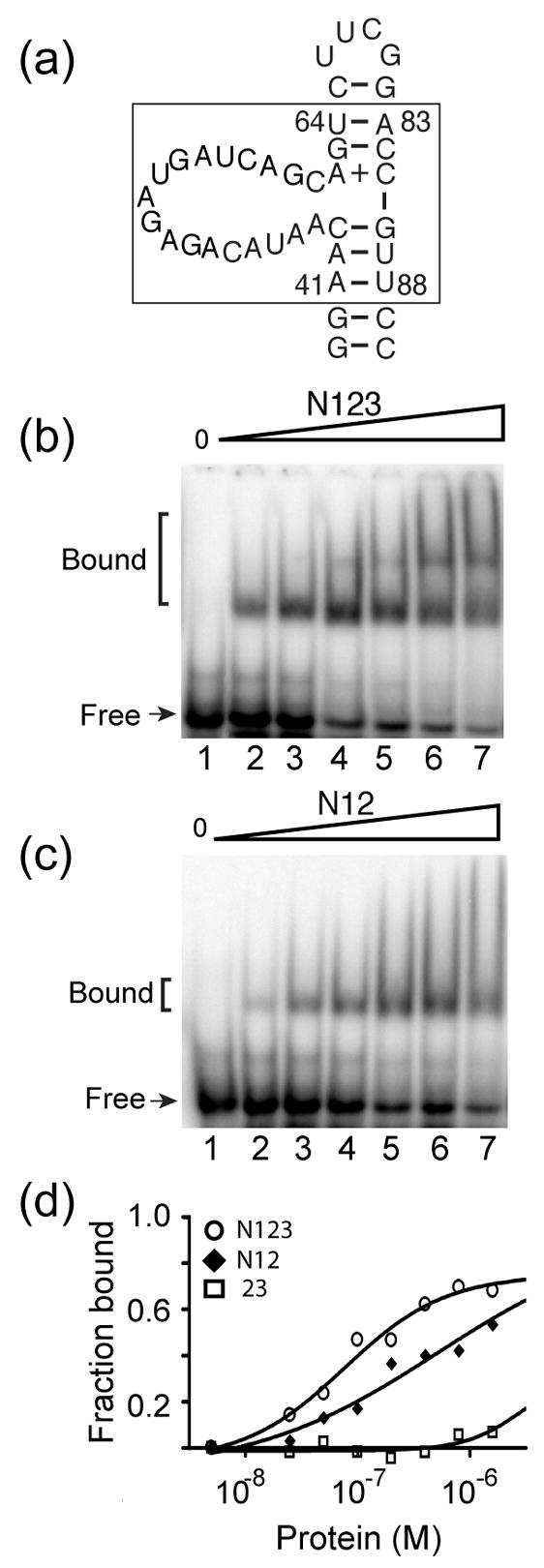
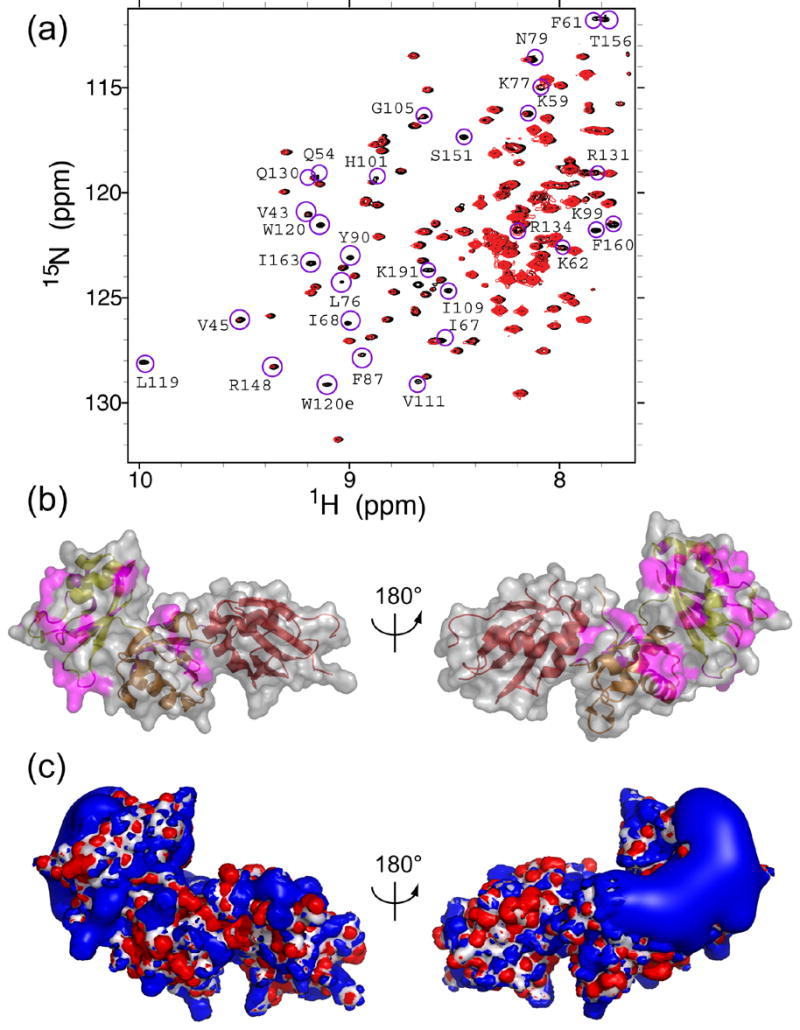
The essential Saccharomyces cerevisiae pre-messenger RNA splicing protein 24 (Prp24) has four RNA recognition motifs (RRMs) and facilitates U6 RNA base-pairing with U4 RNA during spliceosome assembly. Prp24 is a component of the free U6 small nuclear ribonucleoprotein particle (snRNP) but not the U4/U6 bi-snRNP, and so is thought to be displaced from U6 by U4/U6 base-pairing. The interaction partners of each of the four RRMs of Prp24 and how these interactions direct U4/U6 pairing are not known. Here we report the crystal structure of the first three RRMs and the solution structure of the first two RRMs of Prp24. Strikingly, RRM 2 forms extensive inter-domain contacts with RRMs 1 and 3. These contacts occupy much of the canonical RNA-binding faces (beta-sheets) of RRMs 1 and 2, but leave the beta-sheet of RRM 3 exposed. Previously identified substitutions in Prp24 that suppress mutations in U4 and U6 spliceosomal RNAs cluster primarily in the beta-sheet of RRM 3, but also in a conserved loop of RRM 2. RNA binding assays and chemical shift mapping indicate that a large basic patch evident on the surface of RRMs 1 and 2 is part of a high affinity U6 RNA binding site. Our results suggest that Prp24 binds free U6 RNA primarily with RRMs 1 and 2, which may remodel the U6 secondary structure. The beta-sheet of RRM 3 then influences U4/U6 pairing through interaction with an unidentified ligand.
Figures
References
-
- Will CL, Lührmann R. Spliceosome structure and function. In: Gesteland RF, Cech TR, Atkins JF, editors. The RNA World. 3. Cold Spring Harbor Laboratory Press; Cold Spring Harbor, NY: 2006. pp. 369–400.
-
- Staley JP, Guthrie C. Mechanical devices of the spliceosome: motors, clocks, springs, and things. Cell. 1998;92:315–26. - PubMed
-
- Brow DA. Allosteric cascade of spliceosome activation. Annu Rev Genet. 2002;36:333–60. - PubMed
-
- Herschlag D. RNA chaperones and the RNA folding problem. J Biol Chem. 1995;270:20871–4. - PubMed
-
- Schroeder R, Barta A, Semrad K. Strategies for RNA folding and assembly. Nat Rev Mol Cell Biol. 2004;5:908–19. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- P41 RR002301/RR/NCRR NIH HHS/United States
- P50 GM064598/GM/NIGMS NIH HHS/United States
- GM64598/GM/NIGMS NIH HHS/United States
- GM08293/GM/NIGMS NIH HHS/United States
- P41 GM066326/GM/NIGMS NIH HHS/United States
- T32 GM008293/GM/NIGMS NIH HHS/United States
- R01 GM065166/GM/NIGMS NIH HHS/United States
- GM08349/GM/NIGMS NIH HHS/United States
- T32 GM008349/GM/NIGMS NIH HHS/United States
- GM74901/GM/NIGMS NIH HHS/United States
- GM65166/GM/NIGMS NIH HHS/United States
- P41RR02301/RR/NCRR NIH HHS/United States
- GM54018/GM/NIGMS NIH HHS/United States
- P41GM66326/GM/NIGMS NIH HHS/United States
- U54 GM074901/GM/NIGMS NIH HHS/United States
- S10 RR002781/RR/NCRR NIH HHS/United States
- R01 GM054018/GM/NIGMS NIH HHS/United States
- S10 RR008438/RR/NCRR NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
