

Glycolipid transfer proteins
- PMID: 17320476
- PMCID: PMC1986823
- DOI: 10.1016/j.bbalip.2007.01.011
Glycolipid transfer proteins
Abstract
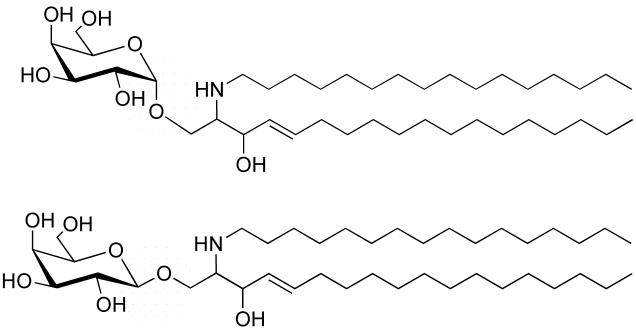
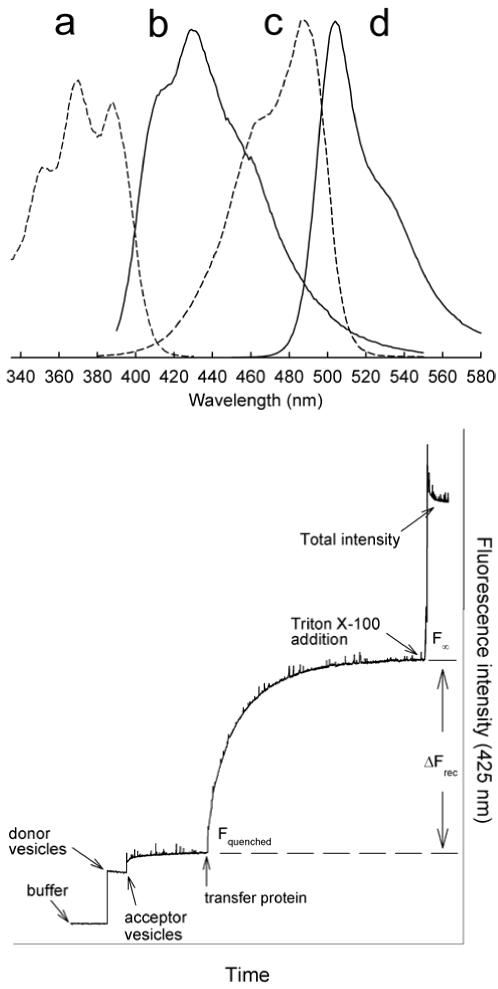
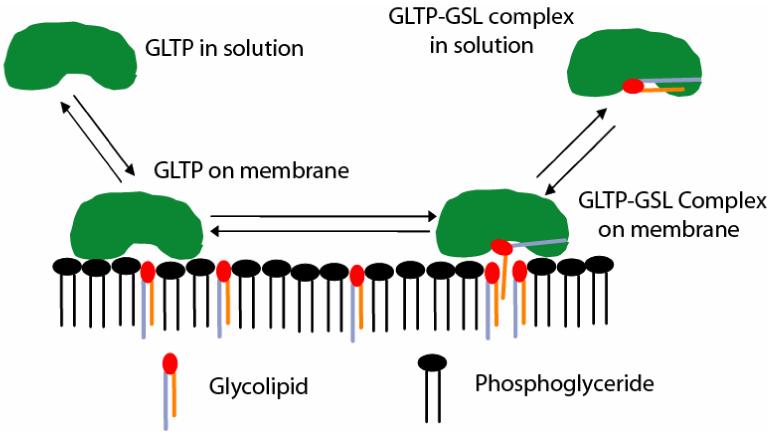
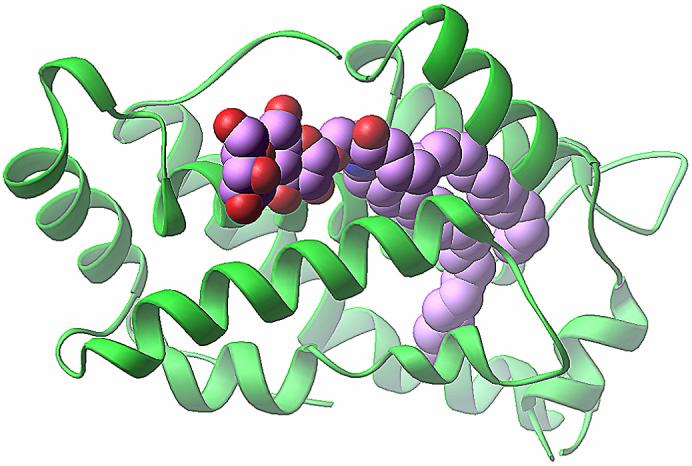
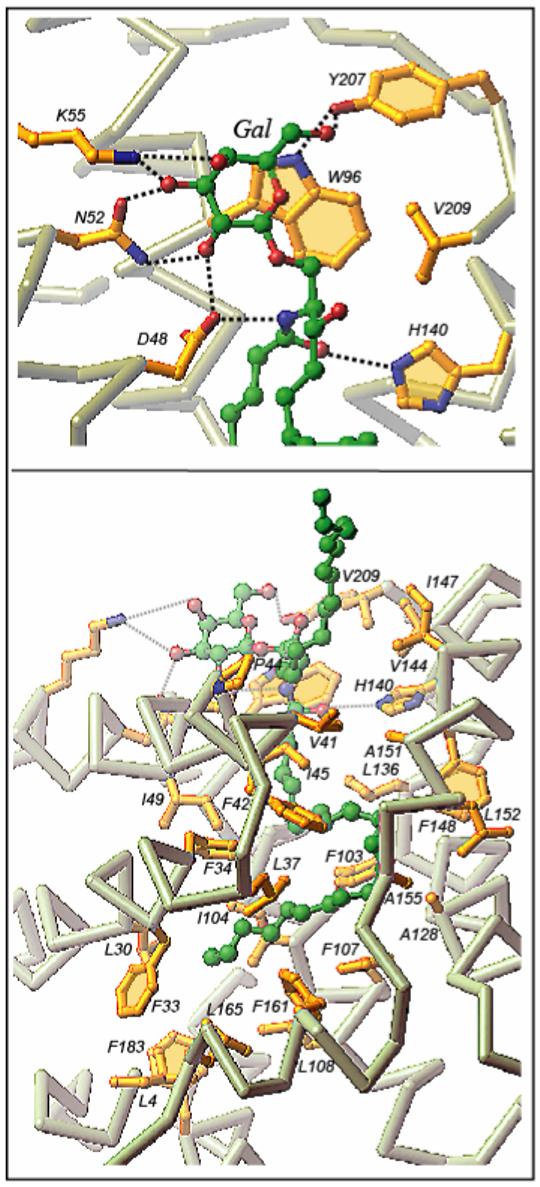
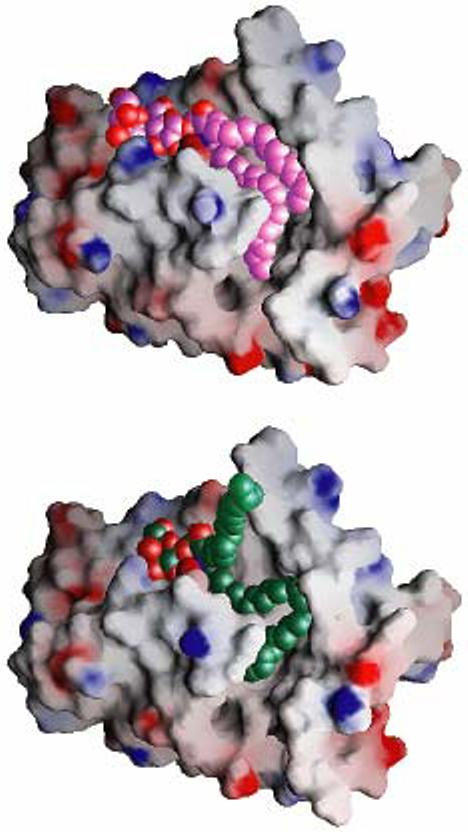
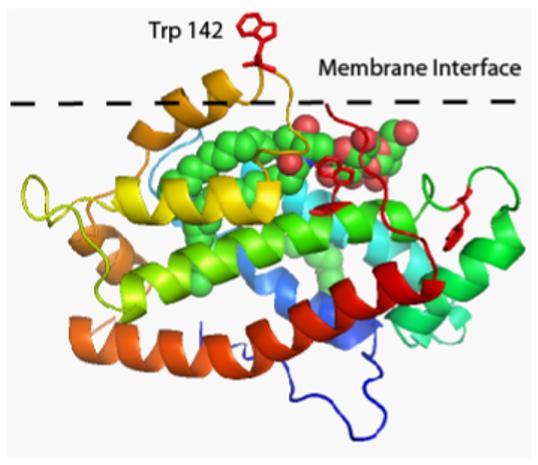
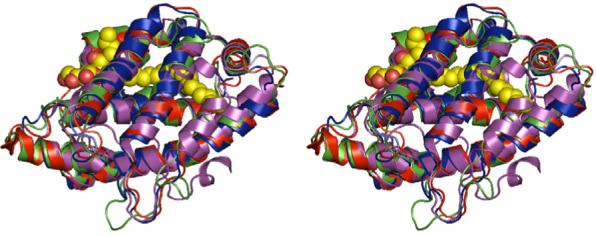
Glycolipid transfer proteins (GLTPs) are small (24 kDa), soluble, ubiquitous proteins characterized by their ability to accelerate the intermembrane transfer of glycolipids in vitro. GLTP specificity encompasses both sphingoid- and glycerol-based glycolipids, but with a strict requirement that the initial sugar residue be beta-linked to the hydrophobic lipid backbone. The 3D architecture of GLTP reveals liganded structures with unique lipid-binding modes. The biochemical properties of GLTP action at the membrane surface have been studied rather comprehensively, but the biological role of GLTP remains enigmatic. What is clear is that GLTP differs distinctly from other known glycolipid-binding proteins, such as nonspecific lipid transfer proteins, lysosomal sphingolipid activator proteins, lectins, lung surfactant proteins as well as other lipid-binding/transfer proteins. Based on the unique conformational architecture that targets GLTP to membranes and enables glycolipid binding, GLTP is now considered the prototypical and founding member of a new protein superfamily in eukaryotes.
Figures
References
-
- Metz RJ, Radin NS. Glucosylceramide uptake from spleen cytosol. J. Biol. Chem. 1980;255:4463–4467. - PubMed
-
- Metz RJ, Radin NS. Purification and properties of a cerebroside transfer protein. J. Biol. Chem. 1982;257:12901–12907. - PubMed
-
- Abe A, Yamada K, Sasaki T. A protein purified from pig brain accelerates the intermembranous translocation of mono- and dihexosylceramides, but not the translocation of phospholipids. Biochem. Biophys. Res. Commun. 1982;104:1386–1393. - PubMed
-
- Wong M, Brown RE, Barenholz Y, Thompson TE. Glycolipid transfer protein from bovine brain. Biochemistry. 1984;23:6498–6505. - PubMed
-
- Abe A, Sasaki T. Purification and some properties of the glycolipid transfer protein from pig brain. J. Biol. Chem. 1985;260:11231–11239. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
