

Mre11 and Ku regulation of double-strand break repair by gene conversion and break-induced replication
- PMID: 17321803
- PMCID: PMC1948817
- DOI: 10.1016/j.dnarep.2007.01.006
Mre11 and Ku regulation of double-strand break repair by gene conversion and break-induced replication
Abstract
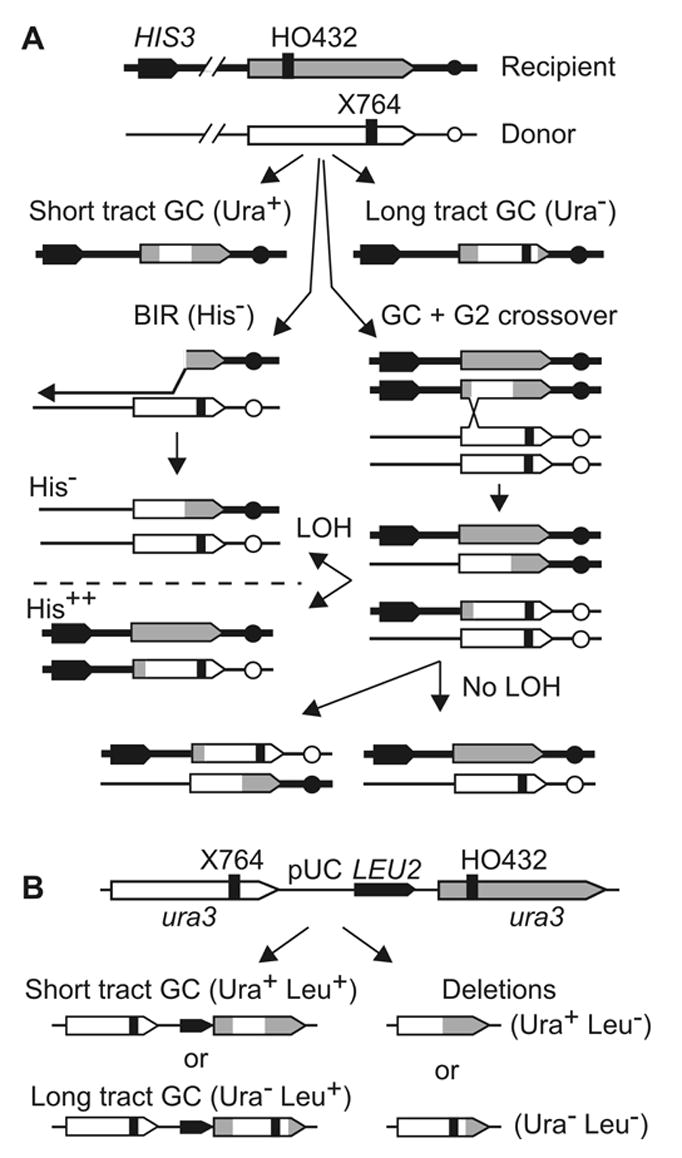
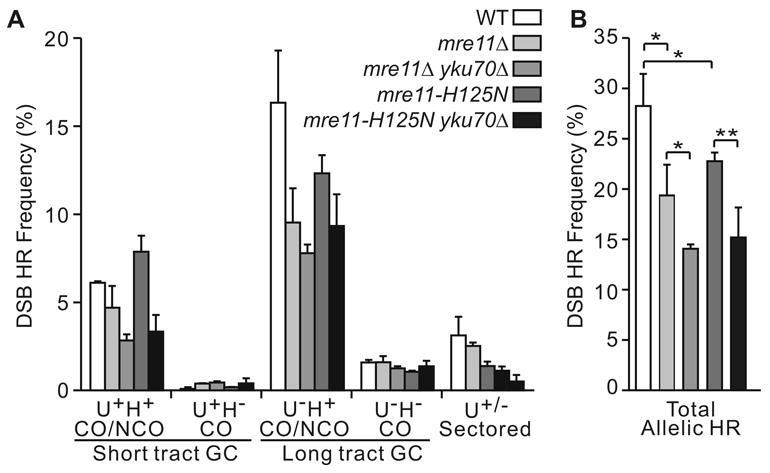
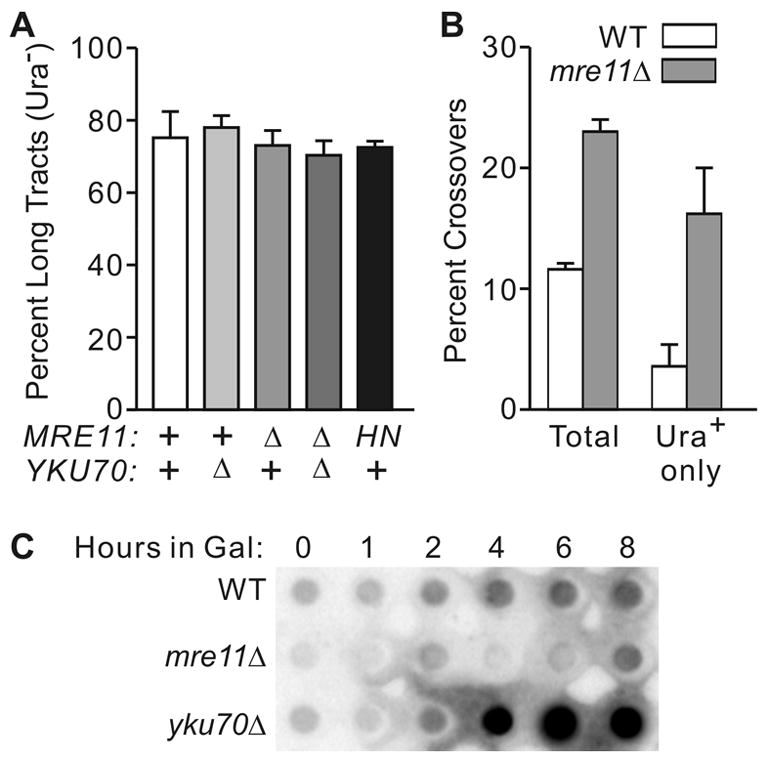
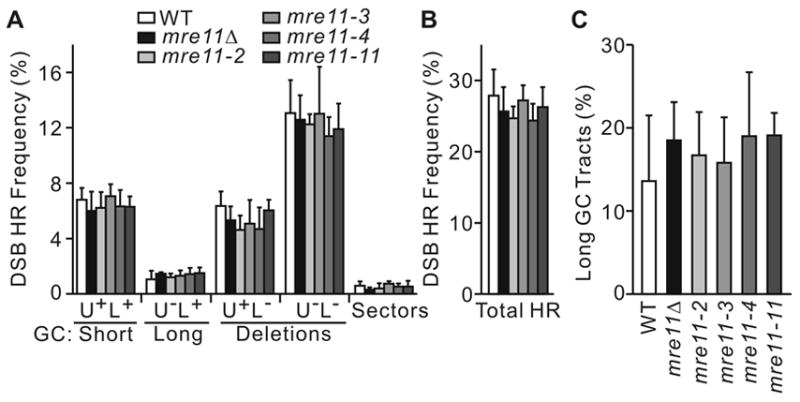
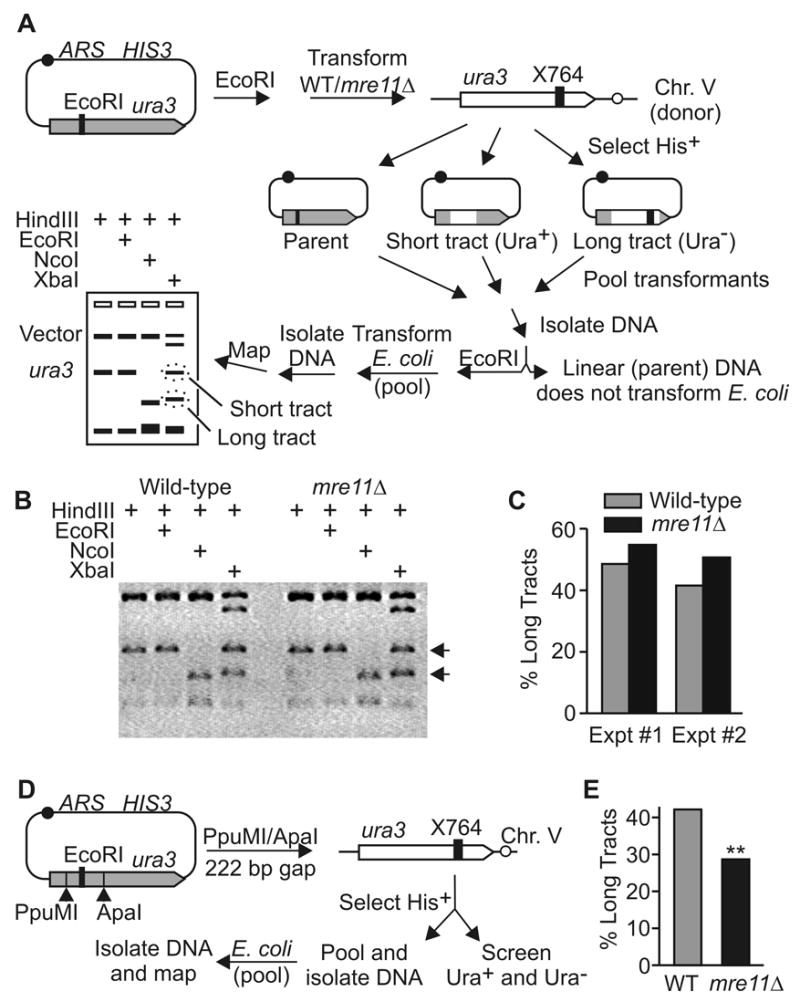
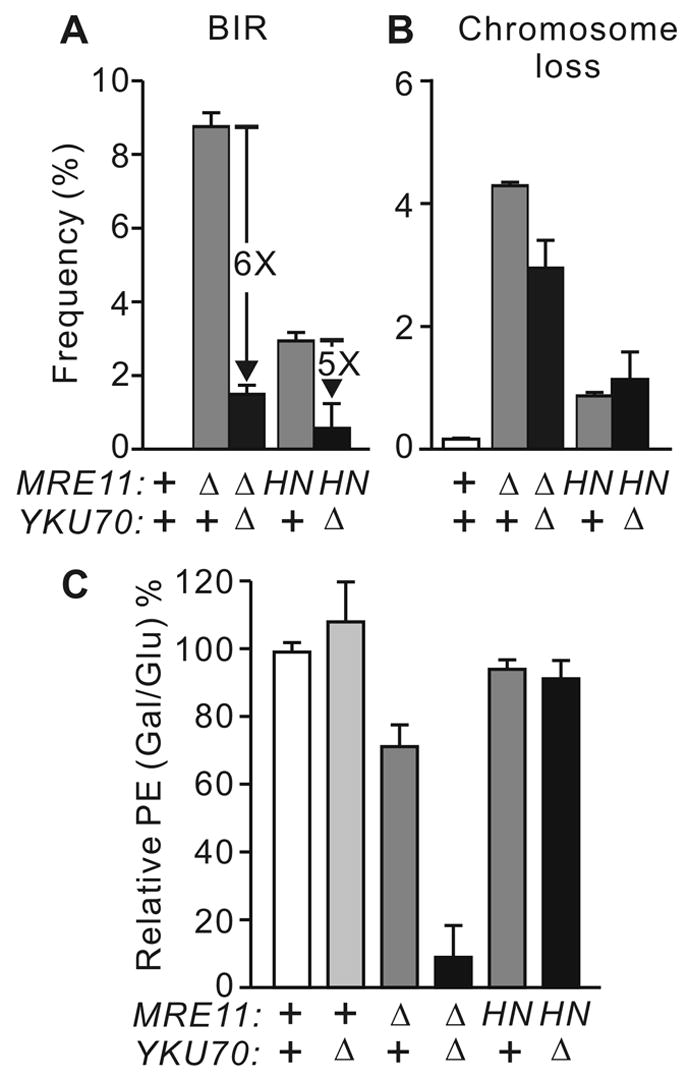
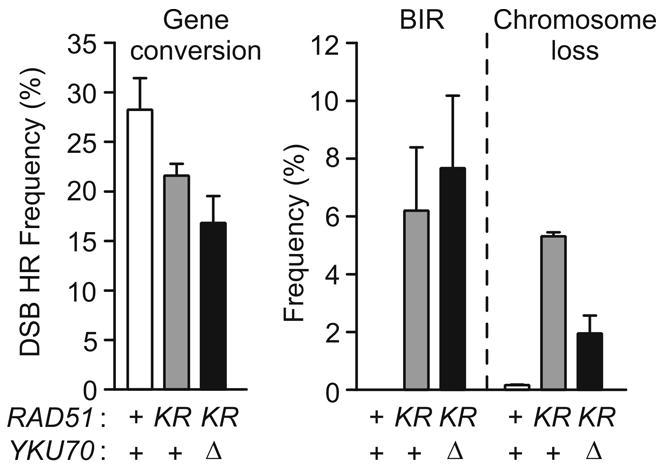
The yeast Mre11-Rad50-Xrs2 (MRX) and Ku complexes regulate single-strand resection at DNA double-strand breaks (DSB), a key early step in homologous recombination (HR). A prior plasmid gap repair study showed that mre11 mutations, which slow single-strand resection, reduce gene conversion tract lengths and the frequency of associated crossovers. Here we tested whether mre11Delta or nuclease-defective mre11 mutations reduced gene conversion tract lengths during HR between homologous chromosomes in diploid yeast. We found that mre11 mutations reduced the efficiency of HR but did not reduce tract lengths or crossovers, despite substantially reduced end-resection at the test (ura3) locus. End-resection is increased in yku70Delta, but this change also had no effect on tract lengths. Thus, heteroduplex formation and tract lengths are not regulated by the extent of end-resection during DSB repair in a chromosomal context. In a plasmid-chromosome DSB repair assay, tract lengths were again similar in wild-type and mre11Delta, but they were reduced in mre11Delta in a gap repair assay. These results indicate that tract lengths are not affected by the extent of end processing when broken ends can invade nearby sites, perhaps because MRX coordination of the two broken ends is dispensable when ends invade nearby sites. Although HR outcome was largely unaffected in mre11 mutants, break-induced replication (BIR) and chromosome loss increased, suggesting that Mre11 function in mitotic HR is limited to early HR stages. Interestingly, yku70Delta suppressed BIR in mre11 mutants. BIR is also elevated in rad51 mutants, but yku70Delta did not suppress BIR in a rad51 background. These results indicate that Mre11 functions in Rad51-independent BIR, and that Ku functions in Rad51-dependent BIR.
Figures
Similar articles
-
Overexpression of Rad51 inhibits double-strand break-induced homologous recombination but does not affect gene conversion tract lengths.DNA Repair (Amst). 2005 Jun 8;4(6):687-98. doi: 10.1016/j.dnarep.2005.03.003. DNA Repair (Amst). 2005. PMID: 15878310
-
Differential suppression of DNA repair deficiencies of Yeast rad50, mre11 and xrs2 mutants by EXO1 and TLC1 (the RNA component of telomerase).Genetics. 2002 Jan;160(1):49-62. doi: 10.1093/genetics/160.1.49. Genetics. 2002. PMID: 11805044 Free PMC article.
-
Release of Ku and MRN from DNA ends by Mre11 nuclease activity and Ctp1 is required for homologous recombination repair of double-strand breaks.PLoS Genet. 2011 Sep;7(9):e1002271. doi: 10.1371/journal.pgen.1002271. Epub 2011 Sep 8. PLoS Genet. 2011. PMID: 21931565 Free PMC article.
-
The multiple roles of the Mre11 complex for meiotic recombination.Chromosome Res. 2007;15(5):551-63. doi: 10.1007/s10577-007-1147-9. Chromosome Res. 2007. PMID: 17674145 Review.
-
DNA double-strand break repair from head to tail.Curr Opin Struct Biol. 2002 Feb;12(1):115-22. doi: 10.1016/s0959-440x(02)00297-x. Curr Opin Struct Biol. 2002. PMID: 11839498 Review.
Cited by
-
Chromosomal Translocations in the Parasite Leishmania by a MRE11/RAD50-Independent Microhomology-Mediated End Joining Mechanism.PLoS Genet. 2016 Jun 17;12(6):e1006117. doi: 10.1371/journal.pgen.1006117. eCollection 2016 Jun. PLoS Genet. 2016. PMID: 27314941 Free PMC article.
-
Lactobacillus sp. Facilitate the Repair of DNA Damage Caused by Bile-Induced Reactive Oxygen Species in Experimental Models of Gastroesophageal Reflux Disease.Antioxidants (Basel). 2023 Jun 21;12(7):1314. doi: 10.3390/antiox12071314. Antioxidants (Basel). 2023. PMID: 37507854 Free PMC article.
-
Competing roles of DNA end resection and non-homologous end joining functions in the repair of replication-born double-strand breaks by sister-chromatid recombination.Nucleic Acids Res. 2013 Feb 1;41(3):1669-83. doi: 10.1093/nar/gks1274. Epub 2012 Dec 18. Nucleic Acids Res. 2013. PMID: 23254329 Free PMC article.
-
A genetic screen for increased loss of heterozygosity in Saccharomyces cerevisiae.Genetics. 2008 Jul;179(3):1179-95. doi: 10.1534/genetics.108.089250. Epub 2008 Jun 18. Genetics. 2008. PMID: 18562670 Free PMC article.
-
Quantitation of DNA double-strand break resection intermediates in human cells.Nucleic Acids Res. 2014 Feb;42(3):e19. doi: 10.1093/nar/gkt1309. Epub 2013 Dec 19. Nucleic Acids Res. 2014. PMID: 24362840 Free PMC article.
References
-
- Nickoloff JA, Haber JE. Mating-type control of DNA repair and recombination in Saccharomyces cerevisiae. In: Nickoloff JA, Hoekstra MF, editors. DNA Damage and Repair, Vol. 3: Advances from Phage to Humans. Humana Press; Totowa, NJ: 2001. pp. 107–124.
-
- Critchlow SE, Jackson SP. DNA end-joining: from yeast to man. Trends Biochem Sci. 1998;23:394–398. - PubMed
-
- Wiltzius JJ, Hohl M, Fleming JC, Petrini JH. The Rad50 hook domain is a critical determinant of Mre11 complex functions. Nat Struct Mol Biol. 2005;12:403–407. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
