

The ftsA* gain-of-function allele of Escherichia coli and its effects on the stability and dynamics of the Z ring
- PMID: 17322202
- PMCID: PMC4757590
- DOI: 10.1099/mic.0.2006/001834-0
The ftsA* gain-of-function allele of Escherichia coli and its effects on the stability and dynamics of the Z ring
Abstract
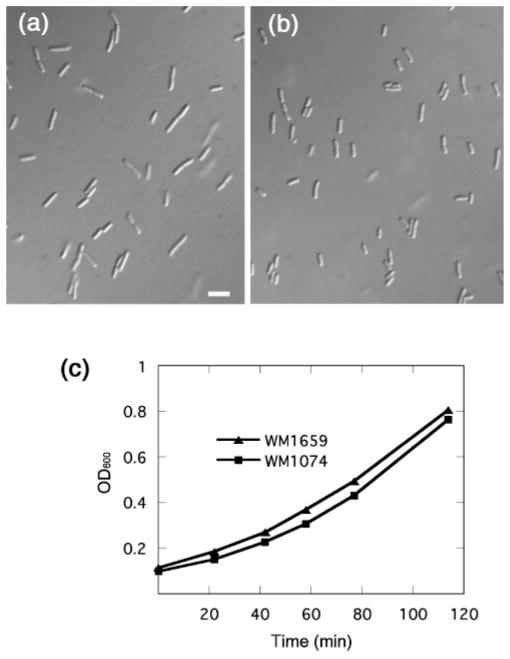
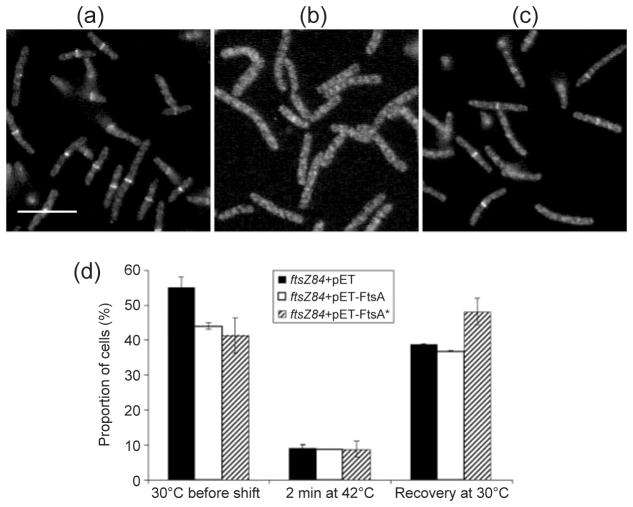
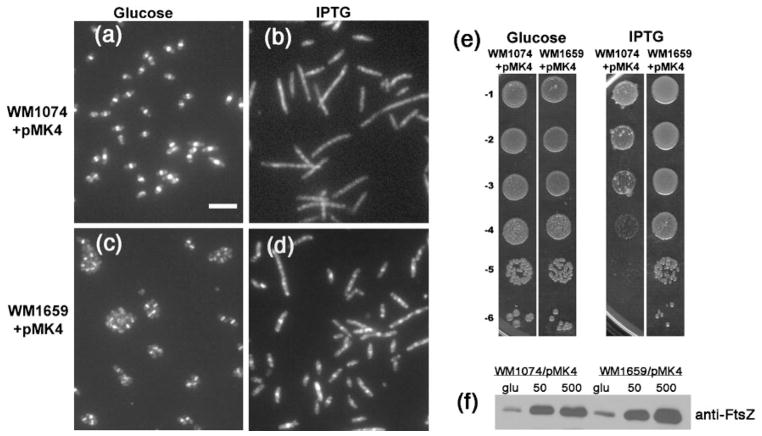
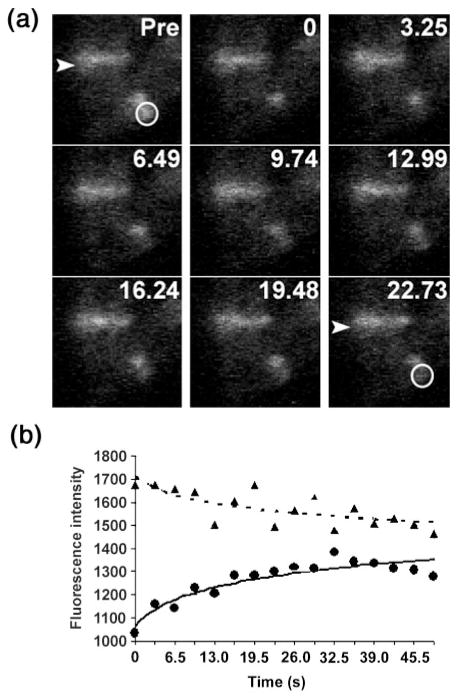
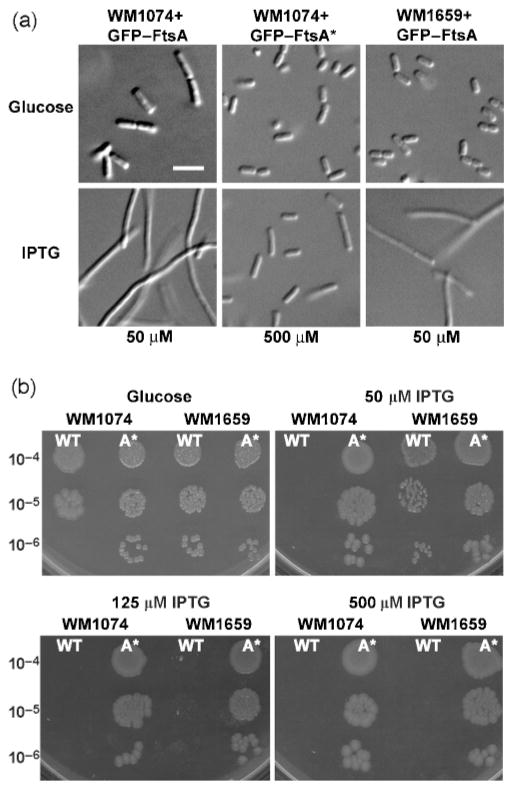
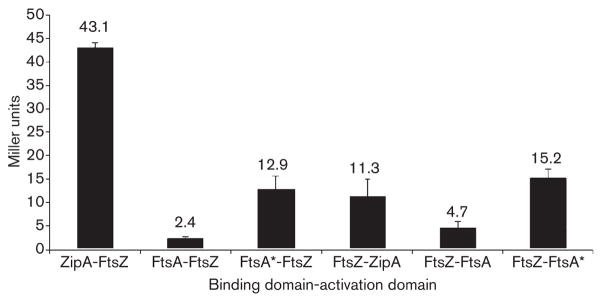
Formation of the FtsZ ring (Z ring) in Escherichia coli is the first step in the assembly of the divisome, a protein machine required for cell division. Although the biochemical functions of most divisome proteins are unknown, several, including ZipA, FtsA and FtsK, have overlapping roles in ensuring that the Z ring assembles at the cytoplasmic membrane, and that it is active. As shown previously, a single amino acid change in FtsA, R286W, also called FtsA*, bypasses the requirement for either ZipA or FtsK in cell division. In this study, the properties of FtsA* were investigated further, with the eventual goal of understanding the molecular mechanism behind the bypass. Compared to wild-type FtsA, the presence of FtsA* resulted in a modest but significant decrease in the mean length of cells in the population, accelerated the reassembly of Z rings, and suppressed the cell-division block caused by excessively high levels of FtsZ. These effects were not mediated by Z-ring remodelling, because FtsA* did not alter the kinetics of FtsZ turnover within the Z ring, as measured by fluorescence recovery after photobleaching. FtsA* was also unable to permit normal cell division at below normal levels of FtsZ, or after thermoinactivation of ftsZ84(ts). However, turnover of FtsA* in the ring was somewhat faster than that of wild-type FtsA, and overexpressed FtsA* did not inhibit cell division as efficiently as wild-type FtsA. Finally, FtsA* interacted more strongly with FtsZ compared with FtsA in a yeast two-hybrid system. These results suggest that FtsA* interacts with FtsZ in a markedly different way compared with FtsA.
Figures
References
-
- Bi E, Lutkenhaus J. FtsZ ring structure associated with division in E. coli. Nature. 1991;354:161–164. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases