

WRKY75 transcription factor is a modulator of phosphate acquisition and root development in Arabidopsis
- PMID: 17322336
- PMCID: PMC1851818
- DOI: 10.1104/pp.106.093971
WRKY75 transcription factor is a modulator of phosphate acquisition and root development in Arabidopsis
Abstract
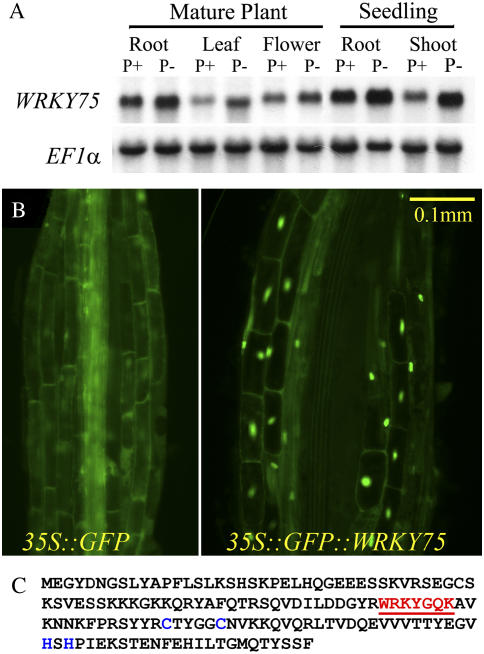
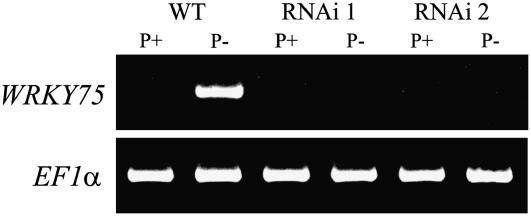
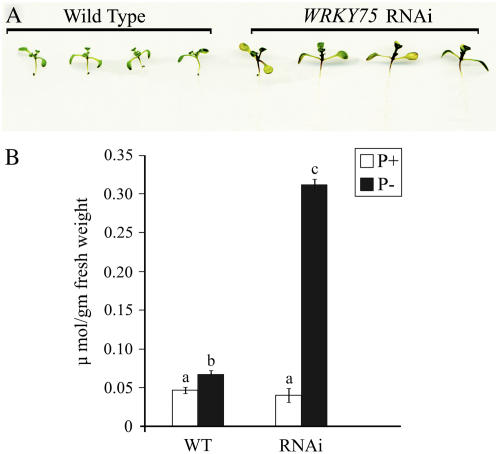
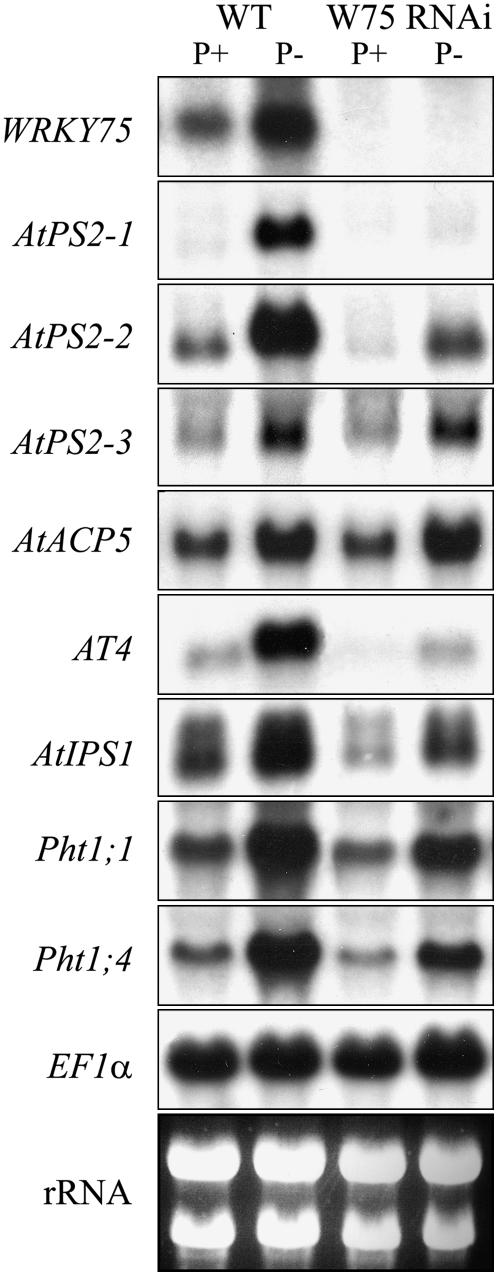
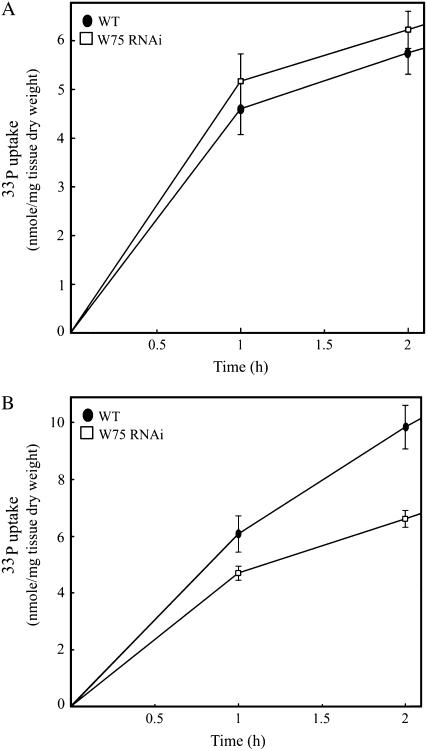
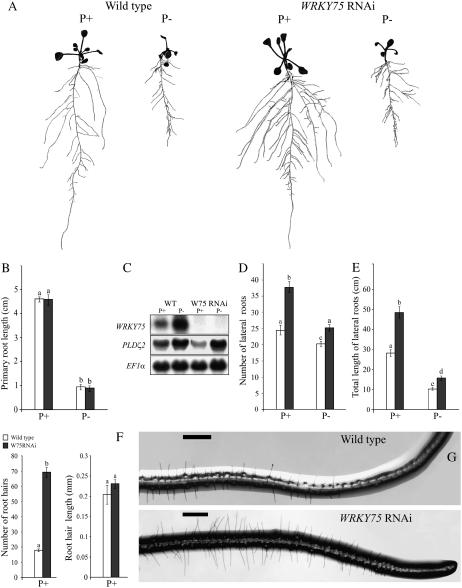
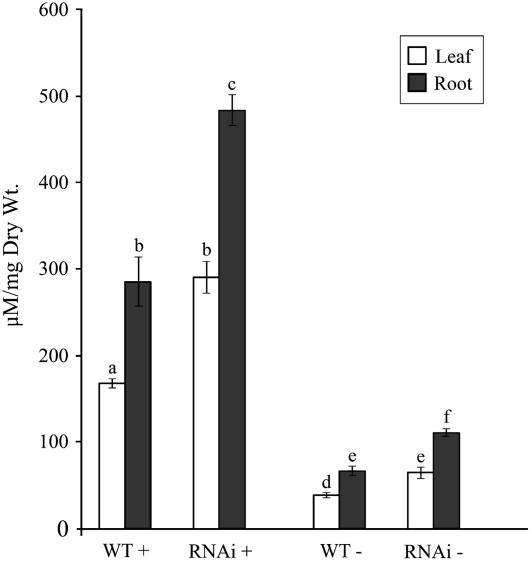
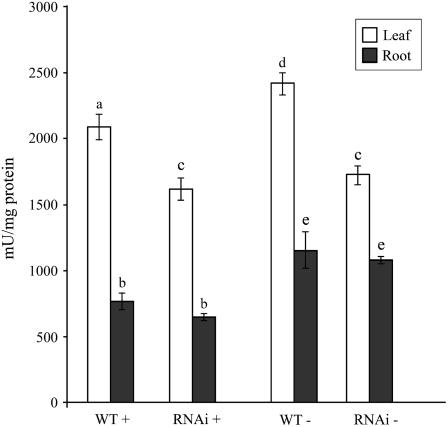
Phosphate (Pi) deficiency limits plant growth and development, resulting in adaptive stress responses. Among the molecular determinants of Pi stress responses, transcription factors play a critical role in regulating adaptive mechanisms. WRKY75 is one of several transcription factors induced during Pi deprivation. In this study, we evaluated the role of the WRKY75 transcription factor in regulating Pi starvation responses in Arabidopsis (Arabidopsis thaliana). WRKY75 was found to be nuclear localized and induced differentially in the plant during Pi deficiency. Suppression of WRKY75 expression through RNAi silencing resulted in early accumulation of anthocyanin, indicating that the RNAi plants were more susceptible to Pi stress. Further analysis revealed that the expression of several genes involved in Pi starvation responses, including phosphatases, Mt4/TPS1-like genes, and high-affinity Pi transporters, was decreased when WRKY75 was suppressed. Consequently, Pi uptake of the mutant plant was also decreased during Pi starvation. In addition, when WRKY75 expression was suppressed, lateral root length and number, as well as root hair number, were significantly increased. However, changes in the root architecture were obvious under both Pi-sufficient and Pi-deficient conditions. This indicates that the regulatory effect of WRKY75 on root architecture could be independent of the Pi status of the plant. Together, these results suggest that WRKY75 is a modulator of Pi starvation responses as well as root development. WRKY75 is the first member of the WRKY transcription factor family reported to be involved in regulating a nutrient starvation response and root development.
Figures
References
-
- Abel S, Ticconi AC, Delatorre CA (2002) Phosphate sensing in higher plants. Physiol Plant 115 1–8 - PubMed
-
- Abramoff MD, Magelhaes PJ, Ram SJ (2004) Image processing with ImageJ. Biophotonics International 11 36–42
-
- Ascencio J (1994) Acid phosphatase as a diagnostic tool. Commun Soil Sci Plant Anal 25 1553–1564
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
