

Basic helix-loop-helix transcription factor heterocomplex of Yas1p and Yas2p regulates cytochrome P450 expression in response to alkanes in the yeast Yarrowia lipolytica
- PMID: 17322346
- PMCID: PMC1865651
- DOI: 10.1128/EC.00412-06
Basic helix-loop-helix transcription factor heterocomplex of Yas1p and Yas2p regulates cytochrome P450 expression in response to alkanes in the yeast Yarrowia lipolytica
Abstract
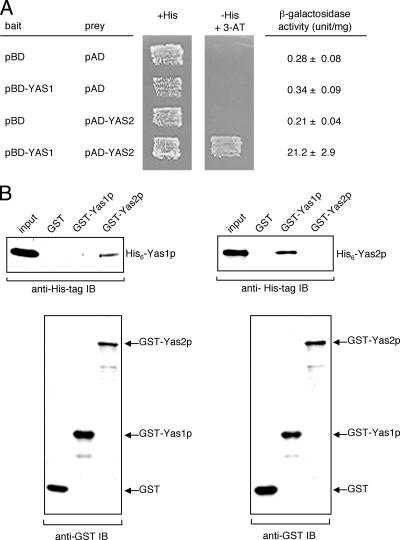
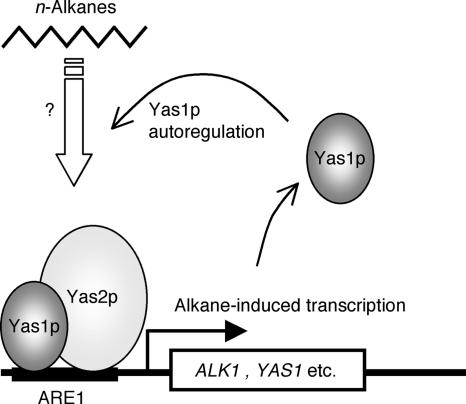
The expression of the ALK1 gene, which encodes cytochrome P450, catalyzing the first step of alkane oxidation in the alkane-assimilating yeast Yarrowia lipolytica, is highly regulated and can be induced by alkanes. Previously, we identified a cis-acting element (alkane-responsive element 1 [ARE1]) in the ALK1 promoter. We showed that a basic helix-loop-helix (bHLH) protein, Yas1p, binds to ARE1 in vivo and mediates alkane-dependent transcription induction. Yas1p, however, does not bind to ARE1 by itself in vitro, suggesting that Yas1p requires another bHLH protein partner for its DNA binding, as many bHLH transcription factors function by forming heterodimers. To identify such a binding partner of Yas1p, here we screened open reading frames encoding proteins with the bHLH motif from the Y. lipolytica genome database and identified the YAS2 gene. The deletion of the YAS2 gene abolished the alkane-responsive induction of ALK1 transcription and the growth of the yeast on alkanes. We revealed that Yas2p has transactivation activity. Furthermore, Yas1p and Yas2p formed a protein complex that was required for the binding of these proteins to ARE1. These findings allow us to postulate a model in which bHLH transcription factors Yas1p and Yas2p form a heterocomplex and mediate the transcription induction in response to alkanes.
Figures

Similar articles
-
Yas3p, an Opi1 family transcription factor, regulates cytochrome P450 expression in response to n-alkanes in Yarrowia lipolytica.J Biol Chem. 2009 Mar 13;284(11):7126-37. doi: 10.1074/jbc.M806864200. Epub 2009 Jan 8. J Biol Chem. 2009. PMID: 19131334 Free PMC article.
-
A basic helix-loop-helix transcription factor essential for cytochrome p450 induction in response to alkanes in yeast Yarrowia lipolytica.J Biol Chem. 2004 May 21;279(21):22183-9. doi: 10.1074/jbc.M313313200. Epub 2004 Mar 24. J Biol Chem. 2004. PMID: 15044482
-
Phosphatidic acid and phosphoinositides facilitate liposome association of Yas3p and potentiate derepression of ARE1 (alkane-responsive element one)-mediated transcription control.Fungal Genet Biol. 2013 Dec;61:100-10. doi: 10.1016/j.fgb.2013.09.008. Epub 2013 Oct 9. Fungal Genet Biol. 2013. PMID: 24120453
-
Metabolism of hydrophobic carbon sources and regulation of it in n-alkane-assimilating yeast Yarrowia lipolytica.Biosci Biotechnol Biochem. 2013;77(6):1149-54. doi: 10.1271/bbb.130164. Epub 2013 Jun 7. Biosci Biotechnol Biochem. 2013. PMID: 23748781 Review.
-
SURVEY AND SUMMARY: Saccharomyces cerevisiae basic helix-loop-helix proteins regulate diverse biological processes.Nucleic Acids Res. 2000 Apr 1;28(7):1499-505. doi: 10.1093/nar/28.7.1499. Nucleic Acids Res. 2000. PMID: 10710415 Free PMC article. Review.
Cited by
-
Phosphatidylserine synthase plays a critical role in the utilization of n-alkanes in the yeast Yarrowia lipolytica.FEMS Yeast Res. 2024 Jan 9;24:foae030. doi: 10.1093/femsyr/foae030. FEMS Yeast Res. 2024. PMID: 39293814 Free PMC article.
-
Development of a promising microbial platform for the production of dicarboxylic acids from biorenewable resources.Biotechnol Biofuels. 2018 Nov 9;11:310. doi: 10.1186/s13068-018-1310-x. eCollection 2018. Biotechnol Biofuels. 2018. PMID: 30455739 Free PMC article.
-
Yas3p, an Opi1 family transcription factor, regulates cytochrome P450 expression in response to n-alkanes in Yarrowia lipolytica.J Biol Chem. 2009 Mar 13;284(11):7126-37. doi: 10.1074/jbc.M806864200. Epub 2009 Jan 8. J Biol Chem. 2009. PMID: 19131334 Free PMC article.
-
The inositol regulon controls viability in Candida glabrata.Microbiology (Reading). 2010 Feb;156(Pt 2):452-462. doi: 10.1099/mic.0.030072-0. Epub 2009 Oct 29. Microbiology (Reading). 2010. PMID: 19875437 Free PMC article.
-
Does co-expression of Yarrowia lipolytica genes encoding Yas1p, Yas2p and Yas3p make a potential alkane-responsive biosensor in Saccharomyces cerevisiae?PLoS One. 2020 Dec 17;15(12):e0239882. doi: 10.1371/journal.pone.0239882. eCollection 2020. PLoS One. 2020. PMID: 33332385 Free PMC article.
References
-
- Ambroziak, J., and S. A. Henry. 1994. INO2 and INO4 gene products, positive regulators of phospholipid biosynthesis in Saccharomyces cerevisiae, form a complex that binds to the INO1 promoter. J. Biol. Chem. 269:15344-15349. - PubMed
-
- Bachhawat, N., Q. Ouyang, and S. A. Henry. 1995. Functional characterization of an inositol-sensitive upstream activation sequence in yeast. A cis-regulatory element responsible for inositol-choline mediated regulation of phospholipid biosynthesis. J. Biol. Chem. 270:25087-25095. - PubMed
-
- Barth, G., and C. Gaillardin. 1997. Physiology and genetics of the dimorphic fungus Yarrowia lipolytica. FEMS Microbiol. Rev. 19:219-237. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
