

Hyperproduction of hyaluronan in neu-induced mammary tumor accelerates angiogenesis through stromal cell recruitment: possible involvement of versican/PG-M
- PMID: 17322391
- PMCID: PMC1864876
- DOI: 10.2353/ajpath.2007.060793
Hyperproduction of hyaluronan in neu-induced mammary tumor accelerates angiogenesis through stromal cell recruitment: possible involvement of versican/PG-M
Abstract
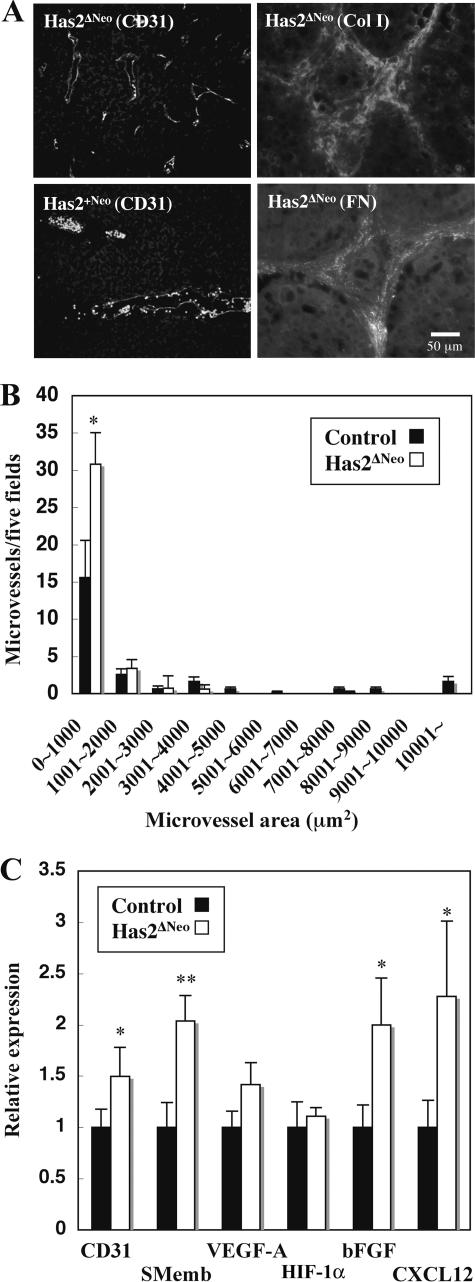
Elevated concentrations of hyaluronan are often associated with human breast cancer malignancy. Here, we investigated the roles of hyaluronan in carcinogenesis and cancer progression using the mouse mammary tumor virus (MMTV)-Neu transgenic model of spontaneous breast cancer. Conditional transgenic mice that express murine hyaluronan synthase 2 (Has2) by Cre-mediated recombination were generated and crossed with the MMTV-Neu mice. In expressing Cre recombinase under the control of the MMTV promoter, the bigenic mice bearing Has2 and neu transgenes exhibited a deposition of hyaluronan matrix and aggressive growth of Neu-initiated mammary tumors. Notably, forced expression of Has2 impaired intercellular adhesion machinery and elicited cell survival signals in tumor cells. Concurrent with these alterations of tumor cells, intratumoral stroma and microvessels were markedly induced. To reveal the molecular basis of hyaluronan-mediated neovascularization, various hyaluronan samples were examined for their ability to potentiate in vivo angiogenesis. In Matrigel plug assays, basic fibroblast growth factor-induced neovascularization was elevated in the presence of either hyaluronan oligosaccharides or a hyaluronan aggregate containing versican. Administration of hyaluronan-versican aggregates, but not native hyaluronan alone, promoted stromal cell recruitment concurrently with the infiltration of endothelial cells. Taken together, these results suggest that hyaluronan overproduction accelerates tumor angiogenesis through stromal reaction, notably in the presence of versican.
Figures







Similar articles
-
Hyaluronan deficiency in tumor stroma impairs macrophage trafficking and tumor neovascularization.Cancer Res. 2010 Sep 15;70(18):7073-83. doi: 10.1158/0008-5472.CAN-09-4687. Epub 2010 Sep 7. Cancer Res. 2010. PMID: 20823158
-
Matrix metalloproteinases play an active role in Wnt1-induced mammary tumorigenesis.Cancer Res. 2006 Mar 1;66(5):2691-9. doi: 10.1158/0008-5472.CAN-05-2919. Cancer Res. 2006. PMID: 16510589
-
A direct test of potential roles for beta3 and beta5 integrins in growth and metastasis of murine mammary carcinomas.Cancer Res. 2005 Nov 15;65(22):10324-9. doi: 10.1158/0008-5472.CAN-04-4098. Cancer Res. 2005. PMID: 16288021
-
Altered hyaluronan biosynthesis in cancer progression.Semin Cancer Biol. 2008 Aug;18(4):268-74. doi: 10.1016/j.semcancer.2008.03.006. Epub 2008 Mar 26. Semin Cancer Biol. 2008. PMID: 18450474 Review.
-
Hyaluronan in human malignancies.Exp Cell Res. 2011 Feb 15;317(4):383-91. doi: 10.1016/j.yexcr.2010.11.017. Epub 2010 Dec 3. Exp Cell Res. 2011. PMID: 21134368 Review.
Cited by
-
TAZ expression as a prognostic indicator in colorectal cancer.PLoS One. 2013;8(1):e54211. doi: 10.1371/journal.pone.0054211. Epub 2013 Jan 23. PLoS One. 2013. PMID: 23372686 Free PMC article.
-
Tumor-Promoting Desmoplasia Is Disrupted by Depleting FAP-Expressing Stromal Cells.Cancer Res. 2015 Jul 15;75(14):2800-2810. doi: 10.1158/0008-5472.CAN-14-3041. Epub 2015 May 15. Cancer Res. 2015. PMID: 25979873 Free PMC article.
-
The anti-inflammatory effects of matrix metalloproteinase-3 on irreversible pulpitis of mature erupted teeth.PLoS One. 2012;7(12):e52523. doi: 10.1371/journal.pone.0052523. Epub 2012 Dec 20. PLoS One. 2012. PMID: 23285075 Free PMC article.
-
Overexpression of hyaluronan-binding protein 1 (HABP1/p32/gC1qR) in HepG2 cells leads to increased hyaluronan synthesis and cell proliferation by up-regulation of cyclin D1 in AKT-dependent pathway.J Biol Chem. 2012 Jun 1;287(23):19750-64. doi: 10.1074/jbc.M111.266270. Epub 2012 Mar 26. J Biol Chem. 2012. PMID: 22451658 Free PMC article.
-
Influence of Hyaluronic Acid Transitions in Tumor Microenvironment on Glioblastoma Malignancy and Invasive Behavior.Front Mater. 2018 Jun;5:39. doi: 10.3389/fmats.2018.00039. Epub 2018 Jun 26. Front Mater. 2018. PMID: 30581816 Free PMC article.
References
-
- Condon MS. The role of the stromal microenvironment in prostate cancer. Semin Cancer Biol. 2005;15:132–137. - PubMed
-
- Mueller MM, Fusenig NE. Friends or foes: bipolar effects of the tumour stroma in cancer. Nat Rev Cancer. 2004;4:839–849. - PubMed
-
- Desmoulière A, Guyot C, Gabbiani G. The stroma reaction myofibroblast: a key player in the control of tumor cell behavior. Int J Dev Biol. 2004;48:509–517. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
