

A core transcriptional network for early mesoderm development in Drosophila melanogaster
- PMID: 17322403
- PMCID: PMC1804332
- DOI: 10.1101/gad.1509007
A core transcriptional network for early mesoderm development in Drosophila melanogaster
Abstract
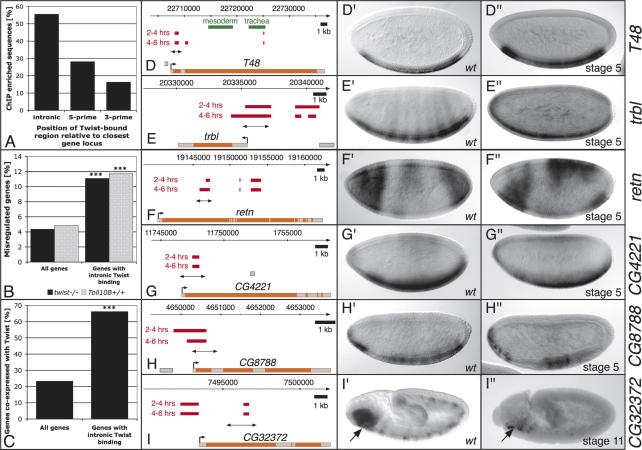
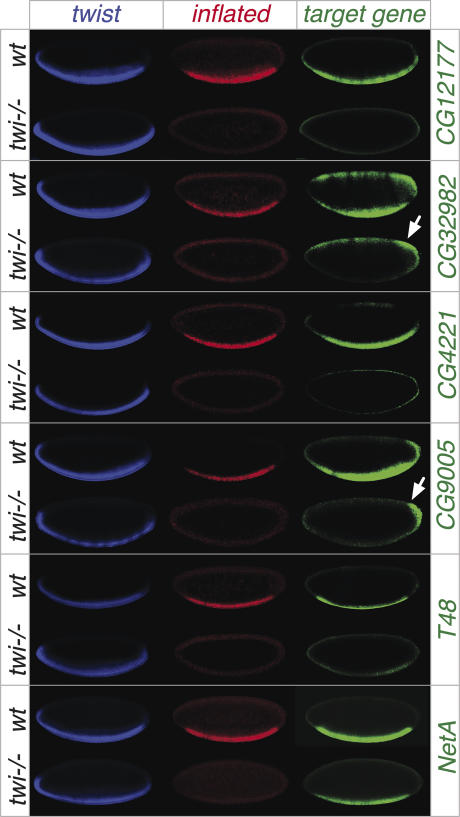
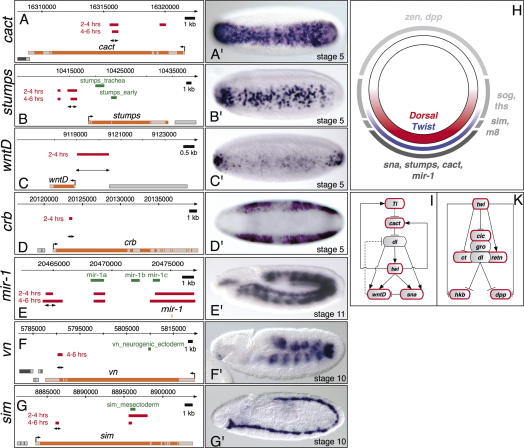
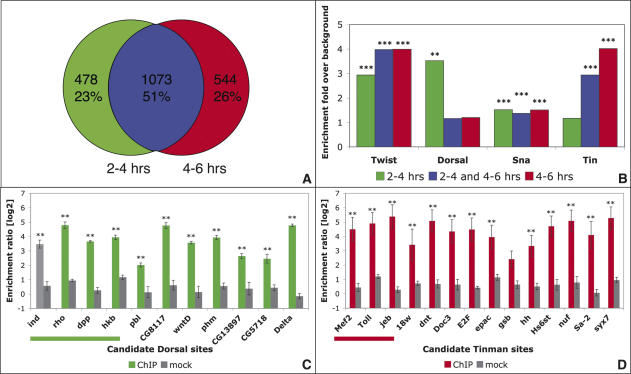
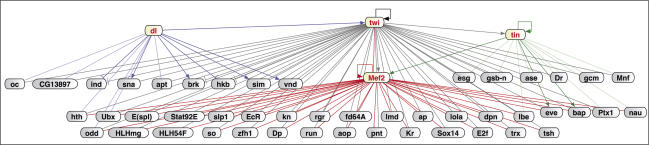
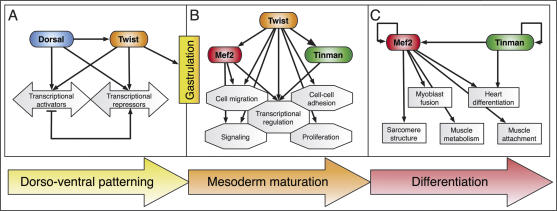
Embryogenesis is controlled by large gene-regulatory networks, which generate spatially and temporally refined patterns of gene expression. Here, we report the characteristics of the regulatory network orchestrating early mesodermal development in the fruitfly Drosophila, where the transcription factor Twist is both necessary and sufficient to drive development. Through the integration of chromatin immunoprecipitation followed by microarray analysis (ChIP-on-chip) experiments during discrete time periods with computational approaches, we identified >2000 Twist-bound cis-regulatory modules (CRMs) and almost 500 direct target genes. Unexpectedly, Twist regulates an almost complete cassette of genes required for cell proliferation in addition to genes essential for morophogenesis and cell migration. Twist targets almost 25% of all annotated Drosophila transcription factors, which may represent the entire set of regulators necessary for the early development of this system. By combining in vivo binding data from Twist, Mef2, Tinman, and Dorsal we have constructed an initial transcriptional network of early mesoderm development. The network topology reveals extensive combinatorial binding, feed-forward regulation, and complex logical outputs as prevalent features. In addition to binary activation and repression, we suggest that Twist binds to almost all mesodermal CRMs to provide the competence to integrate inputs from more specialized transcription factors.
Figures

References
-
- Arora K., Nusslein-Volhard C., Nusslein-Volhard C. Altered mitotic domains reveal fate map changes in Drosophila embryos mutant for zygotic dorsoventral patterning genes. Development. 1992;114:1003–1024. - PubMed
-
- Barolo S., Carver L.A., Posakony J.W., Carver L.A., Posakony J.W., Posakony J.W. GFP and β-galactosidase transformation vectors for promoter/enhancer analysis in Drosophila. Biotechniques. 2000;29:726, 728, 730, 732. - PubMed
-
- Bate M., Rushton E., Currie D.A., Rushton E., Currie D.A., Currie D.A. Cells with persistent twist expression are the embryonic precursors of adult muscles in Drosophila. Development. 1991;113:79–89. - PubMed
-
- Baylies M.K., Bate M., Bate M. twist: A myogenic switch in Drosophila. Science. 1996;272:1481–1484. - PubMed
-
- Beer J., Technau G.M., Campos-Ortega J.A., Technau G.M., Campos-Ortega J.A., Campos-Ortega J.A. Lineage analysis of transplanted individual cells in embryos of Drosophila melanogaster. Roux’s Arch. Dev. Biol. 1987;196:222–230. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases