

Centromere function and nondisjunction are independent components of the maize B chromosome accumulation mechanism
- PMID: 17322406
- PMCID: PMC1867328
- DOI: 10.1105/tpc.106.049577
Centromere function and nondisjunction are independent components of the maize B chromosome accumulation mechanism
Abstract
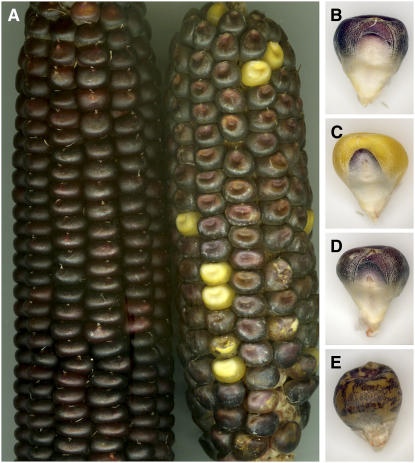
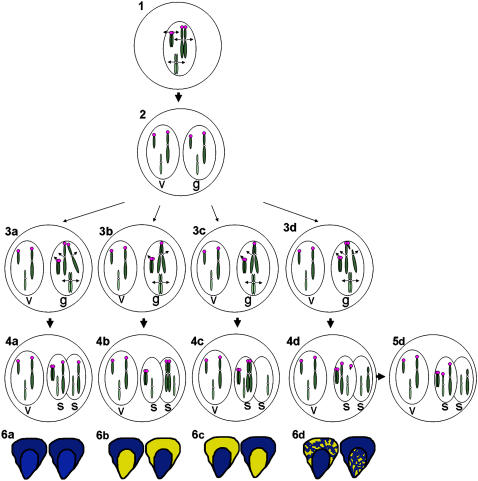
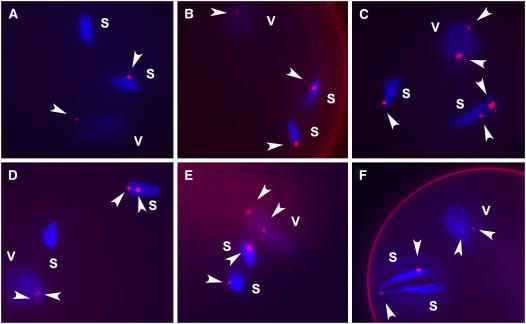
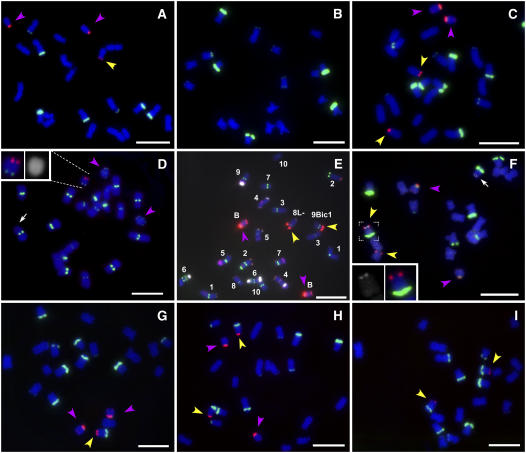
Supernumerary or B chromosomes are selfish entities that maintain themselves in populations by accumulation mechanisms. The accumulation mechanism of the B chromosome of maize (Zea mays) involves nondisjunction at the second pollen mitosis, placing two copies of the B chromosome into one of the two sperm. The B chromosome long arm must be present in the same nucleus for the centromere to undergo nondisjunction. A centromere, containing all of the normal DNA elements, translocated from the B chromosome to the short arm of chromosome 9 was recently found to be epigenetically silenced for centromeric function. When intact B chromosomes were added to this genotype, thus supplying the long arm, the inactive centromere regained the property of nondisjunction causing the translocation chromosome 9 to be differentially distributed to the two sperm or resulted in chromosome breaks in 9S, occasionally producing new translocations. Translocation of the inactive B centromere to chromosome 7 transferred the nondisjunction property to this chromosome. The results provide insight into the molecular and evolutionary basis of this B chromosome accumulation mechanism by demonstrating that nondisjunction is caused by a process that does not depend on normal centromere function but that the region of the chromosome required for nondisjunction resides in the centromeric region.
Figures
References
-
- Beckett, J.B. (1991). Cytogenetic, genetic and plant breeding applications of B-A translocation in maize. In Chromosome Engineering in Plants: Genetics, Breeding and Evolution, P.K. Gupta and T. Tsuchiya, eds (New York: Elsevier), pp. 493–529.
-
- Burt, A., and Trivers, R. (2006). Genes in Conflict. (Cambridge, MA: Belknap Press).
-
- Carlson, W.R. (1978). The B chromosome of corn. Annu. Rev. Genet. 12 5–23. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
