

A novel multimeric form of FasL modulates the ability of diabetogenic T cells to mediate type 1 diabetes in an adoptive transfer model
- PMID: 17324464
- PMCID: PMC1950258
- DOI: 10.1016/j.molimm.2007.01.014
A novel multimeric form of FasL modulates the ability of diabetogenic T cells to mediate type 1 diabetes in an adoptive transfer model
Abstract
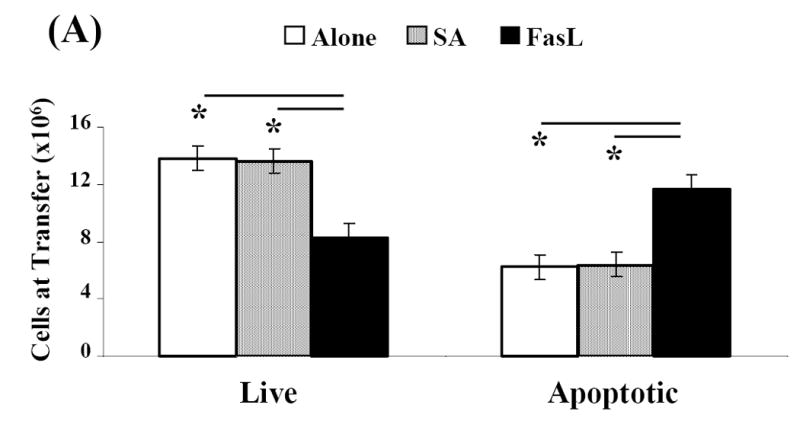
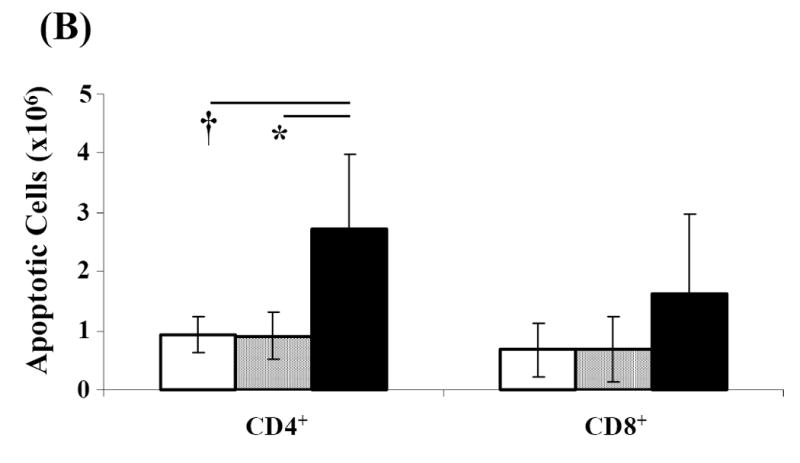
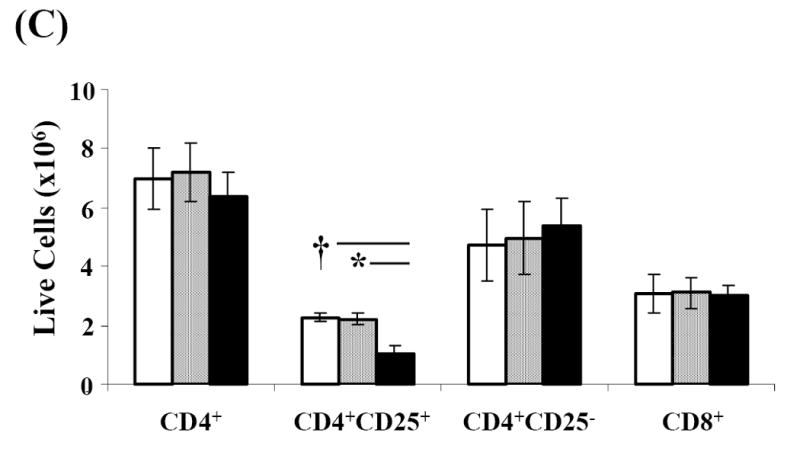
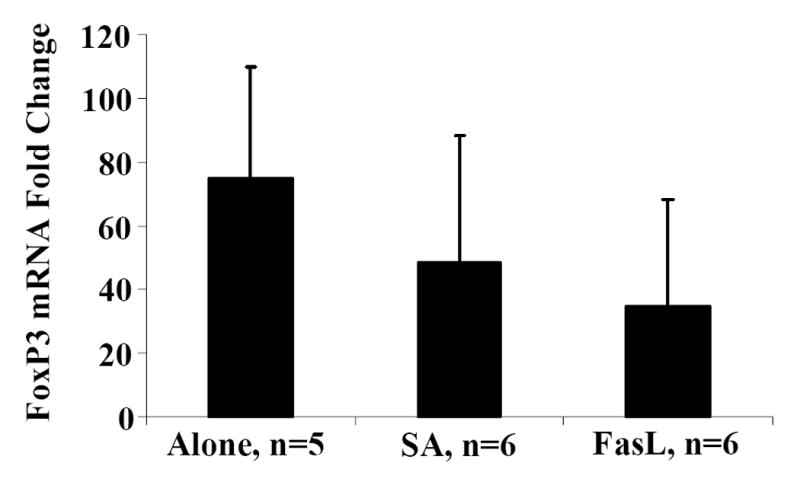
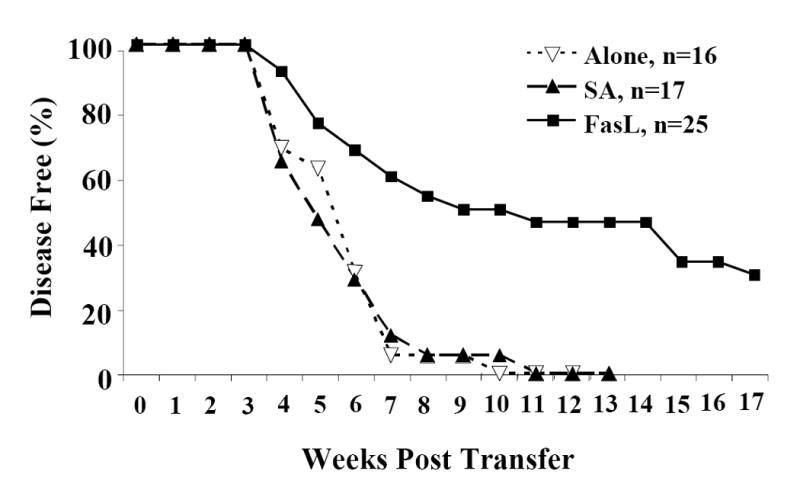
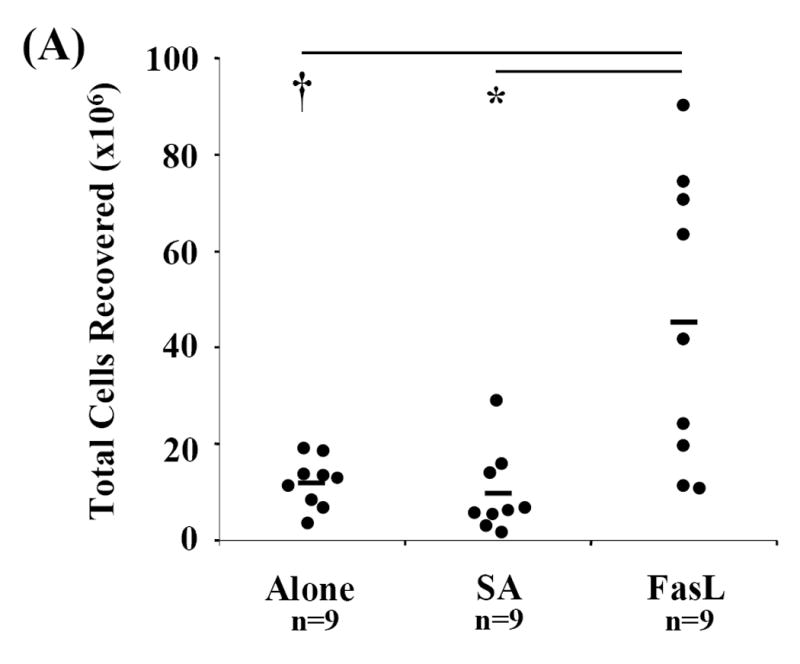
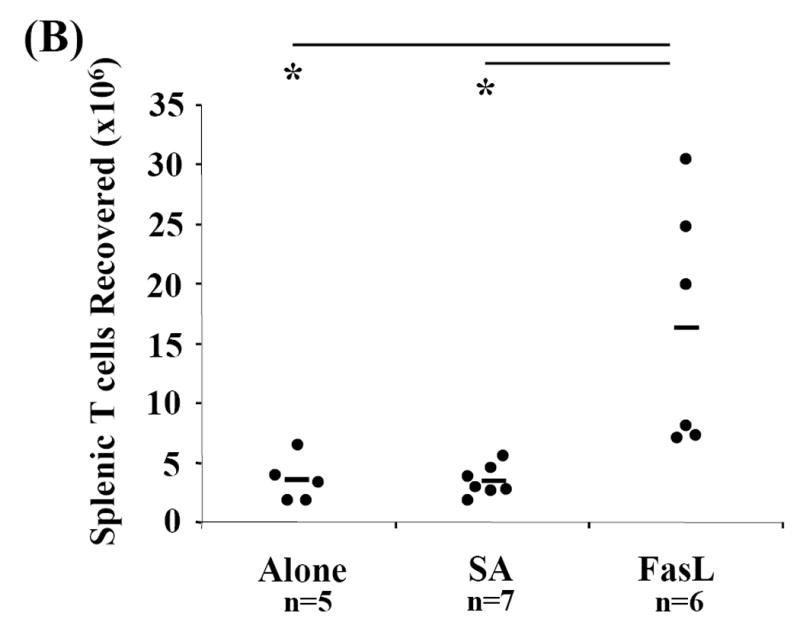
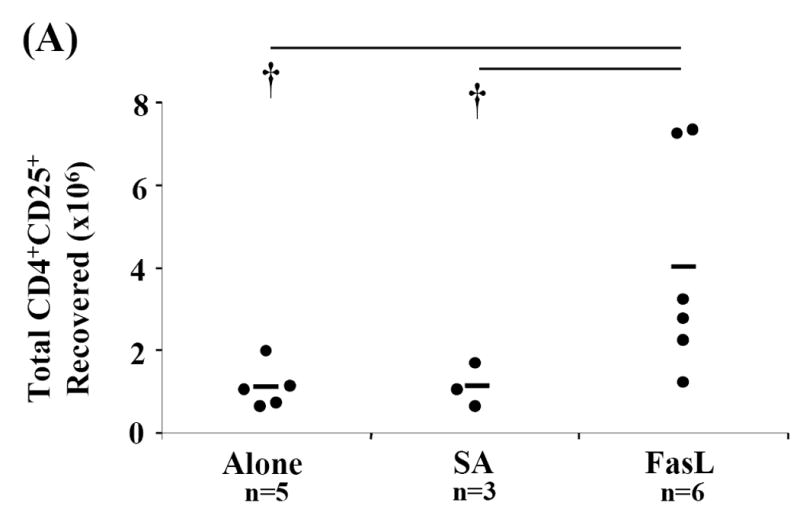
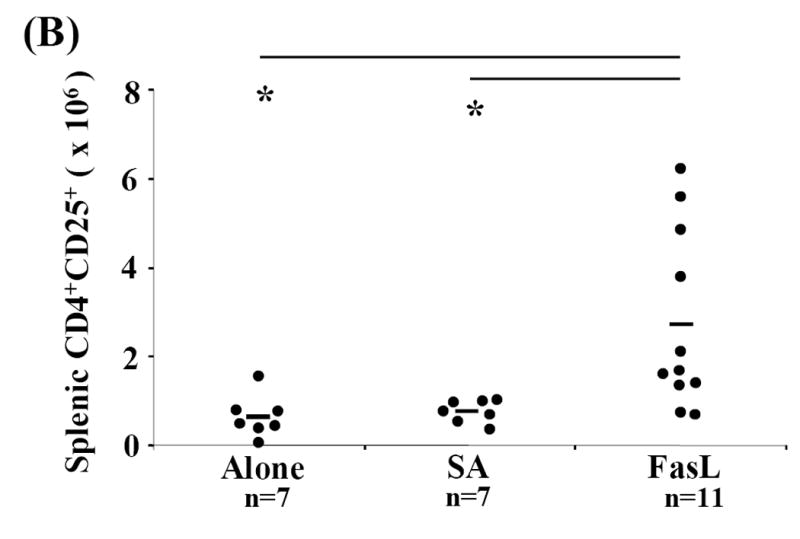
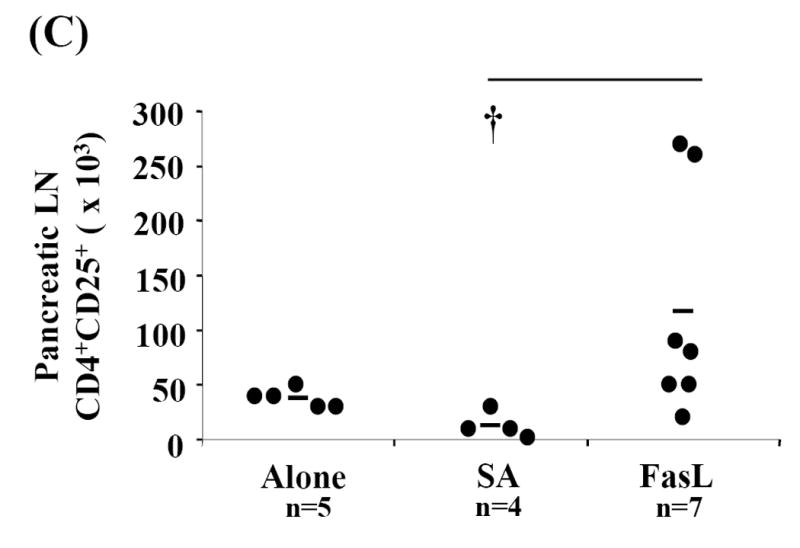
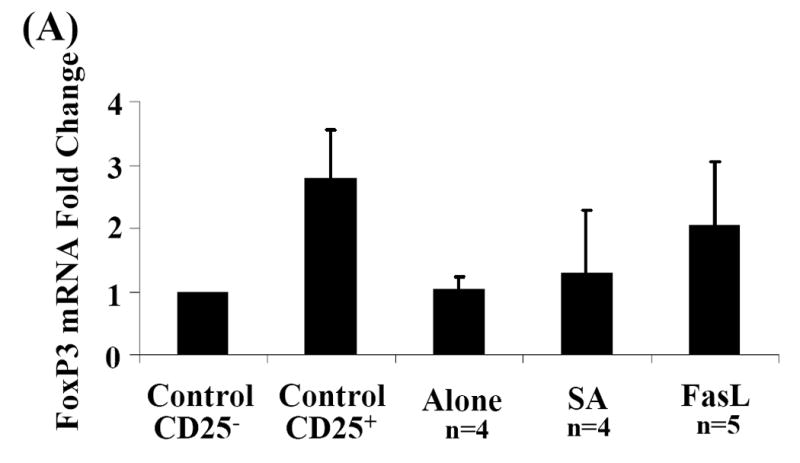
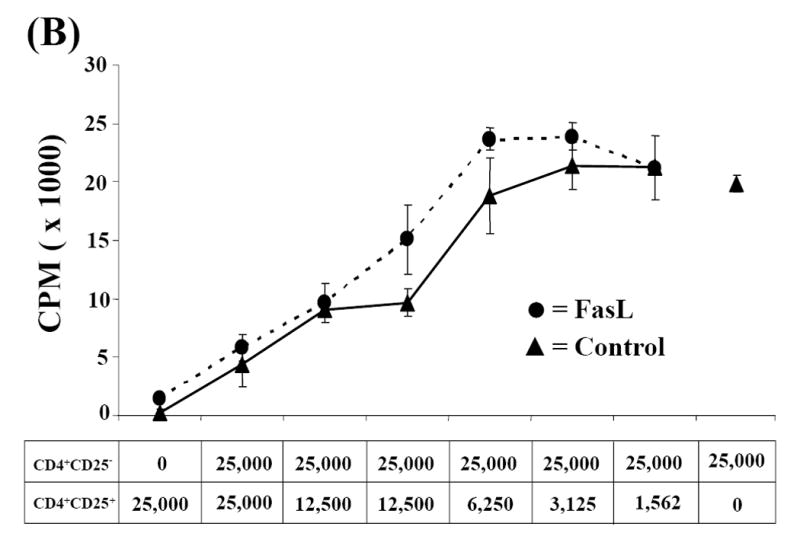
Activation induced cell death (AICD) via Fas/FasL is the primary homeostatic molecular mechanism employed by the immune system to control activated T-cell responses and promote tolerance to self-antigens. We herein investigated the ability of a novel multimeric form of FasL chimeric with streptavidin (SA-FasL) having potent apoptotic activity to induce apoptosis in diabetogenic T cells and modulate insulin-dependent type 1 diabetes (IDDM) in an adoptive transfer model. Diabetogenic splenocytes from NOD/Lt females were co-cultured in vitro with SA-FasL, SA control protein, or alone without protein, and adoptively transferred into NOD/Lt-Rag1(null) recipients for diabetes development. All animals receiving control (Alone: n=16 or SA: n=17) cells developed diabetes on average by 6 weeks, whereas animals receiving SA-FasL-treated (n=25) cells exhibited significantly delayed progression (p<.001) and decreased incidence (70%). This effect was associated with an increase in CD4(+)CD25(+) T cells and correlated with FoxP3 expression in pancreatic lymph nodes. Extracorporeal treatment of peripheral blood lymphocytes using SA-FasL during disease onset represents a novel approach that may alter the ability of pathogenic T cells to mediate diabetes and have therapeutic utility in clinical management of IDDM.
Figures
Similar articles
-
Pancreatic islets engineered with SA-FasL protein establish robust localized tolerance by inducing regulatory T cells in mice.J Immunol. 2011 Dec 1;187(11):5901-9. doi: 10.4049/jimmunol.1003266. Epub 2011 Nov 7. J Immunol. 2011. PMID: 22068235 Free PMC article.
-
Significant role for Fas in the pathogenesis of autoimmune diabetes.J Immunol. 2000 Mar 1;164(5):2523-32. doi: 10.4049/jimmunol.164.5.2523. J Immunol. 2000. PMID: 10679090
-
CD4+CD25+ regulatory T cells generated in response to insulin B:9-23 peptide prevent adoptive transfer of diabetes by diabetogenic T cells.J Autoimmun. 2003 Nov;21(3):221-37. doi: 10.1016/s0896-8411(03)00114-8. J Autoimmun. 2003. PMID: 14599847
-
Inhibition of autoimmune diabetes by Fas ligand: the paradox is solved.J Immunol. 2000 Mar 15;164(6):2931-6. doi: 10.4049/jimmunol.164.6.2931. J Immunol. 2000. PMID: 10706679
-
In vivo diabetogenic action of CD4+ T lymphocytes requires Fas expression and is independent of IL-1 and IL-18.Eur J Immunol. 2011 May;41(5):1344-51. doi: 10.1002/eji.201041216. Epub 2011 Apr 13. Eur J Immunol. 2011. PMID: 21469125 Free PMC article.
Cited by
-
Targeting of IL-2 receptor with a caspase fusion protein disrupts autoimmunity in prediabetic and diabetic NOD mice.Diabetologia. 2010 Feb;53(2):356-68. doi: 10.1007/s00125-009-1604-4. Epub 2009 Nov 28. Diabetologia. 2010. PMID: 19946662
-
Adenovirus-Mediated FasL Minigene Transfer Endows Transduced Cells with Killer Potential.Int J Mol Sci. 2020 Aug 20;21(17):6011. doi: 10.3390/ijms21176011. Int J Mol Sci. 2020. PMID: 32825521 Free PMC article.
-
RGD-avidin-biotin pretargeting to alpha v beta 3 integrin enhances the proapoptotic activity of TNF alpha related apoptosis inducing ligand (TRAIL).Apoptosis. 2008 Feb;13(2):225-35. doi: 10.1007/s10495-007-0166-5. Apoptosis. 2008. PMID: 18071905 Free PMC article.
-
Pancreatic islets engineered with SA-FasL protein establish robust localized tolerance by inducing regulatory T cells in mice.J Immunol. 2011 Dec 1;187(11):5901-9. doi: 10.4049/jimmunol.1003266. Epub 2011 Nov 7. J Immunol. 2011. PMID: 22068235 Free PMC article.
-
Immunomodulation with SA-FasL protein as an effective means of preventing islet allograft rejection in chemically diabetic NOD mice.Transplant Proc. 2013 Jun;45(5):1889-91. doi: 10.1016/j.transproceed.2013.01.041. Transplant Proc. 2013. PMID: 23769064 Free PMC article.
References
-
- Aoki CA, Borchers AT, Ridgway WM, et al. NOD mice and autoimmunity. Autoimmun Rev. 2005;4:373–379. - PubMed
-
- Arreaza G, Salojin K, Yang W, et al. Deficient activation and resistance to activationinduced apoptosis of CD8+ T cells is associated with defective peripheral tolerance in nonobese diabetic mice. Clin Immunol. 2003;107:103–115. - PubMed
-
- Askenasy N, Yolcu ES, Wang Z, et al. Display of Fas ligand protein on cardiac vasculature as a novel means of regulating allograft rejection. Circulation. 2003;107:1525–1531. - PubMed
-
- Askenasy N, Yolcu ES, Yaniv I, et al. Induction of tolerance using Fas ligand: a double-edged immunomodulator. Blood. 2005;105:1396–1404. - PubMed
-
- Bacchetta R, Gregori S, Roncarolo MG. CD4+ regulatory T cells: mechanisms of induction and effector function. Autoimmun Rev. 2005;4:491–496. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
