

Role for non-proteolytic control of M-phase-promoting factor activity at M-phase exit
- PMID: 17327911
- PMCID: PMC1803016
- DOI: 10.1371/journal.pone.0000247
Role for non-proteolytic control of M-phase-promoting factor activity at M-phase exit
Abstract
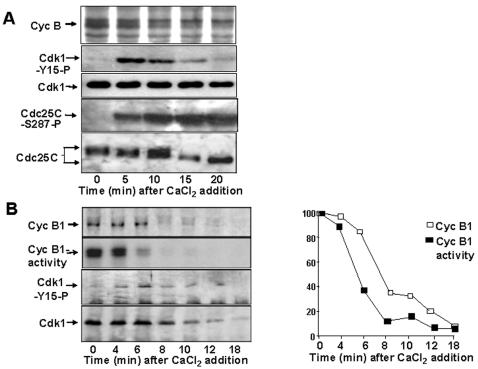
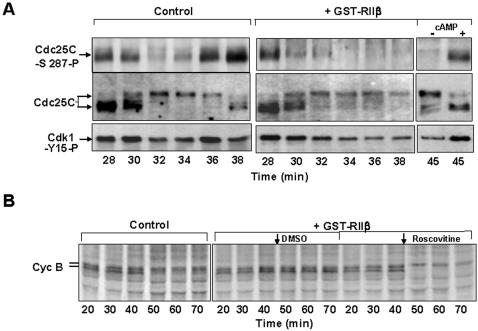
M-phase Promoting Factor (MPF; the cyclin B-cdk 1 complex) is activated at M-phase onset by removal of inhibitory phosphorylation of cdk1 at thr-14 and tyr-15. At M-phase exit, MPF is destroyed by ubiquitin-dependent cyclin proteolysis. Thus, control of MPF activity via inhibitory phosphorylation is believed to be particularly crucial in regulating transition into, rather than out of, M-phase. Using the in vitro cell cycle system derived form Xenopus eggs, here we show, however, that inhibitory phosphorylation of cdk1 contributes to control MPF activity during M-phase exit. By sampling extracts at very short intervals during both meiotic and mitotic exit, we found that cyclin B1-associated cdk1 underwent transient inhibitory phosphorylation at tyr-15 and that cyclin B1-cdk1 activity fell more rapidly than the cyclin B1 content. Inhibitory phosphorylation of MPF correlated with phosphorylation changes of cdc25C, the MPF phosphatase, and physical interaction of cdk1 with wee1, the MPF kinase, during M-phase exit. MPF down-regulation required Ca(++)/calmodulin-dependent kinase II (CaMKII) and cAMP-dependent protein kinase (PKA) activities at meiosis and mitosis exit, respectively. Treatment of M-phase extracts with a mutant cyclin B1-cdk1AF complex, refractory to inhibition by phosphorylation, impaired binding of the Anaphase Promoting Complex/Cyclosome (APC/C) to its co-activator Cdc20 and altered M-phase exit. Thus, timely M-phase exit requires a tight coupling of proteolysis-dependent and proteolysis-independent mechanisms of MPF inactivation.
Conflict of interest statement
Figures



References
-
- Dunphy WG. The decision to enter mitosis. Trends Cell Biol. 1994;4:202–207. - PubMed
-
- Peters JM. The anaphase-promoting complex: proteolysis in mitosis and beyond. Mol Cell. 2002;9:931–943. - PubMed
-
- Yudkovsky Y, Shteinberg M, Listovsky T, Brandeis M, Hershko A. Phosphorylation of Cdc20/fizzy negatively regulates the mammalian cyclosome/APC in the mitotic checkpoint. Biochem Biophys Res Commun. 2000;271:299–304. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
