

The distributed auditory cortex
- PMID: 17329049
- PMCID: PMC2637155
- DOI: 10.1016/j.heares.2007.01.017
The distributed auditory cortex
Abstract
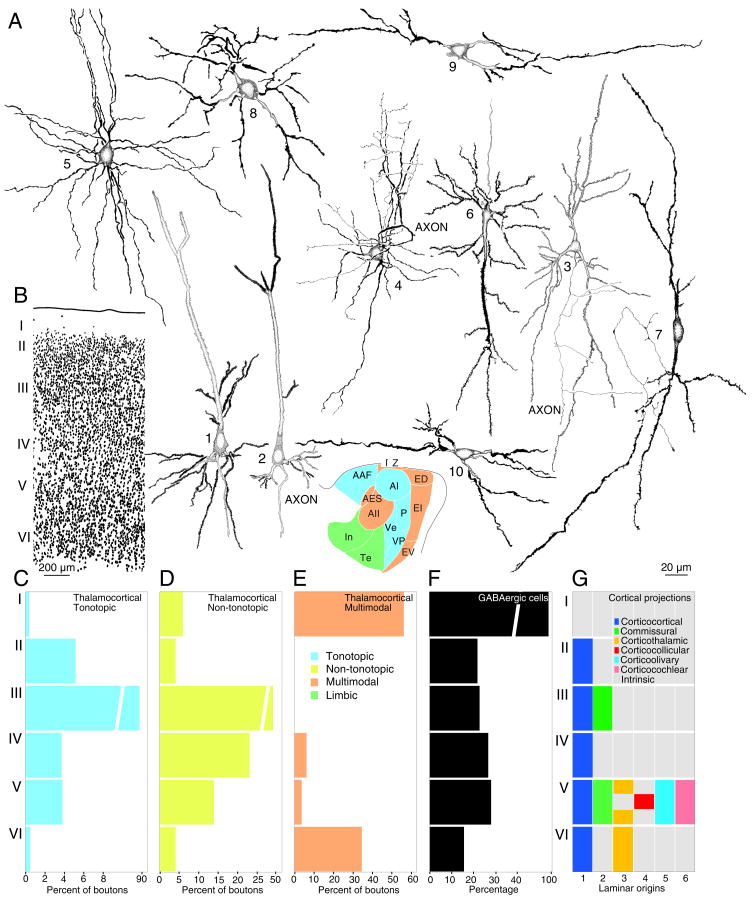
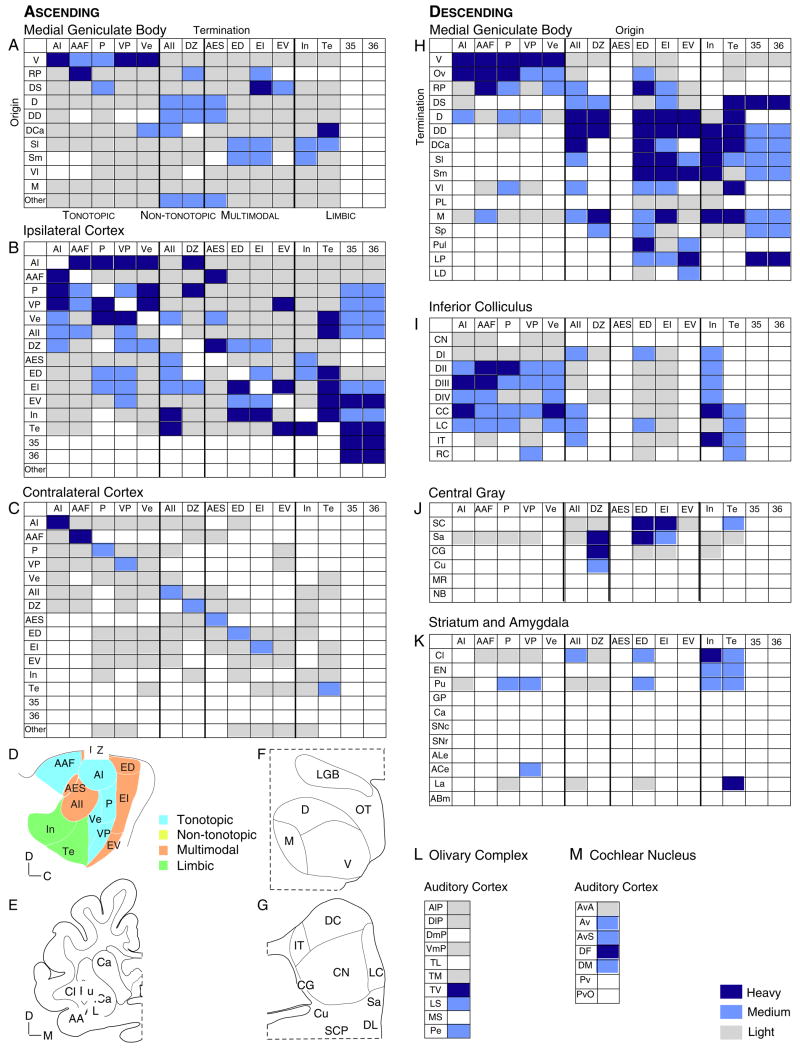
A synthesis of cat auditory cortex (AC) organization is presented in which the extrinsic and intrinsic connections interact to derive a unified profile of the auditory stream and use it to direct and modify cortical and subcortical information flow. Thus, the thalamocortical input provides essential sensory information about peripheral stimulus events, which AC redirects locally for feature extraction, and then conveys to parallel auditory, multisensory, premotor, limbic, and cognitive centers for further analysis. The corticofugal output influences areas as remote as the pons and the cochlear nucleus, structures whose effects upon AC are entirely indirect, and it has diverse roles in the transmission of information through the medial geniculate body and inferior colliculus. The distributed AC is thus construed as a functional network in which the auditory percept is assembled for subsequent redistribution in sensory, premotor, and cognitive streams contingent on the derived interpretation of the acoustic events. The confluence of auditory and multisensory streams likely precedes cognitive processing of sound. The distributed AC constitutes the largest and arguably the most complete representation of the auditory world. Many facets of this scheme may apply in rodent and primate AC as well. We propose that the distributed auditory cortex contributes to local processing regimes in regions as disparate as the frontal pole and the cochlear nucleus to construct the acoustic percept.
Figures

Similar articles
-
Connections of cat auditory cortex: I. Thalamocortical system.J Comp Neurol. 2008 Apr 20;507(6):1879-900. doi: 10.1002/cne.21611. J Comp Neurol. 2008. PMID: 18271026 Free PMC article.
-
Decoding the auditory corticofugal systems.Hear Res. 2005 Sep;207(1-2):1-9. doi: 10.1016/j.heares.2005.06.007. Hear Res. 2005. Corrected and republished in: Hear Res. 2006 Feb;212(1-2):1-8. doi: 10.1016/j.heares.2005.06.014. PMID: 16091301 Corrected and republished. Review.
-
Multisensory processing via early cortical stages: Connections of the primary auditory cortical field with other sensory systems.Neuroscience. 2006 Dec 28;143(4):1065-83. doi: 10.1016/j.neuroscience.2006.08.035. Epub 2006 Oct 4. Neuroscience. 2006. PMID: 17027173
-
Functional architecture of auditory cortex.Curr Opin Neurobiol. 2002 Aug;12(4):433-40. doi: 10.1016/s0959-4388(02)00342-2. Curr Opin Neurobiol. 2002. PMID: 12139992 Review.
-
Convergence of thalamic and cortical pathways in cat auditory cortex.Hear Res. 2011 Apr;274(1-2):85-94. doi: 10.1016/j.heares.2010.05.008. Epub 2010 May 26. Hear Res. 2011. PMID: 20576491 Free PMC article. Review.
Cited by
-
Coding of auditory-stimulus identity in the auditory non-spatial processing stream.J Neurophysiol. 2008 Jan;99(1):87-95. doi: 10.1152/jn.01069.2007. Epub 2007 Nov 14. J Neurophysiol. 2008. PMID: 18003874 Free PMC article.
-
Behavioral detection of intra-cortical microstimulation in the primary and secondary auditory cortex of cats.Front Syst Neurosci. 2015 Apr 27;9:61. doi: 10.3389/fnsys.2015.00061. eCollection 2015. Front Syst Neurosci. 2015. PMID: 25964744 Free PMC article.
-
Precise memory for pure tones is predicted by measures of learning-induced sensory system neurophysiological plasticity at cortical and subcortical levels.Learn Mem. 2020 Jul 15;27(8):328-339. doi: 10.1101/lm.051318.119. Print 2020 Aug. Learn Mem. 2020. PMID: 32669388 Free PMC article.
-
State-dependent encoding of sound and behavioral meaning in a tertiary region of the ferret auditory cortex.Nat Neurosci. 2019 Mar;22(3):447-459. doi: 10.1038/s41593-018-0317-8. Epub 2019 Jan 28. Nat Neurosci. 2019. PMID: 30692690 Free PMC article.
-
When and How Does the Auditory Cortex Influence Subcortical Auditory Structures? New Insights About the Roles of Descending Cortical Projections.Front Neurosci. 2021 Aug 3;15:690223. doi: 10.3389/fnins.2021.690223. eCollection 2021. Front Neurosci. 2021. PMID: 34413722 Free PMC article. Review.
References
-
- Aitkin LM. Medial geniculate body of the cat: responses to tonal stimuli of neurons in medial division. J Neurophysiol. 1973;36:275–283. - PubMed
-
- Aitkin LM, Dunlop CW. Interplay of excitation and inhibition in the cat medial geniculate body. J Neurophysiol. 1968;31:44–61. - PubMed
-
- Bajo VM, Rouiller EM, Welker E, Clarke S, Villa AEP, de Ribaupierre Y, de Ribaupierre F. Morphology and spatial distribution of corticothalamic terminals originating from the cat auditory cortex. Hearing Res. 1995;83:161–174. - PubMed
-
- Bao S, Chan VT, Merzenich MM. Cortical remodelling induced by activity of ventral tegmental dopamine neurons. Nature. 2001;412:79–83. - PubMed
-
- Beneyto M, Prieto JJ. Connections of the auditory cortex with the claustrum and endopiriform nucleus in the cat. Brain Res Bull. 2001;54:485–498. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
