

NMDA receptor function and NMDA receptor-dependent phosphorylation of huntingtin is altered by the endocytic protein HIP1
- PMID: 17329427
- PMCID: PMC6673493
- DOI: 10.1523/JNEUROSCI.5175-06.2007
NMDA receptor function and NMDA receptor-dependent phosphorylation of huntingtin is altered by the endocytic protein HIP1
Abstract
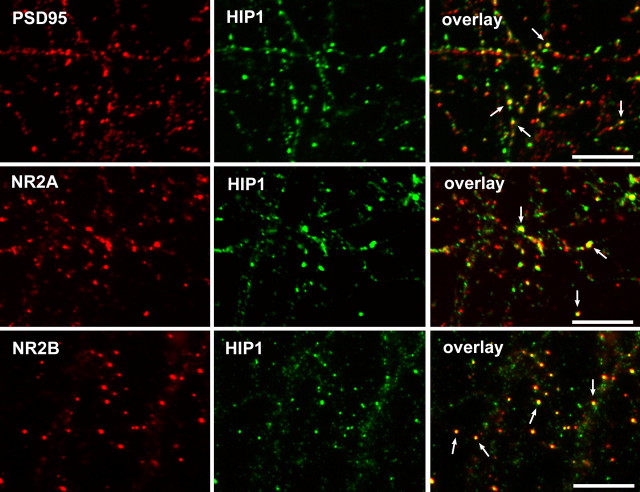
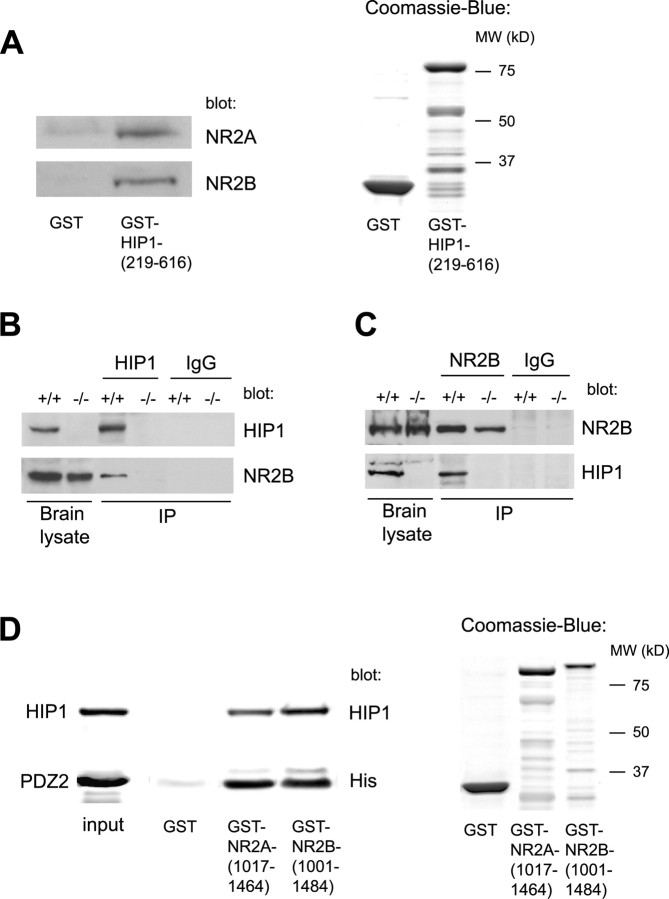
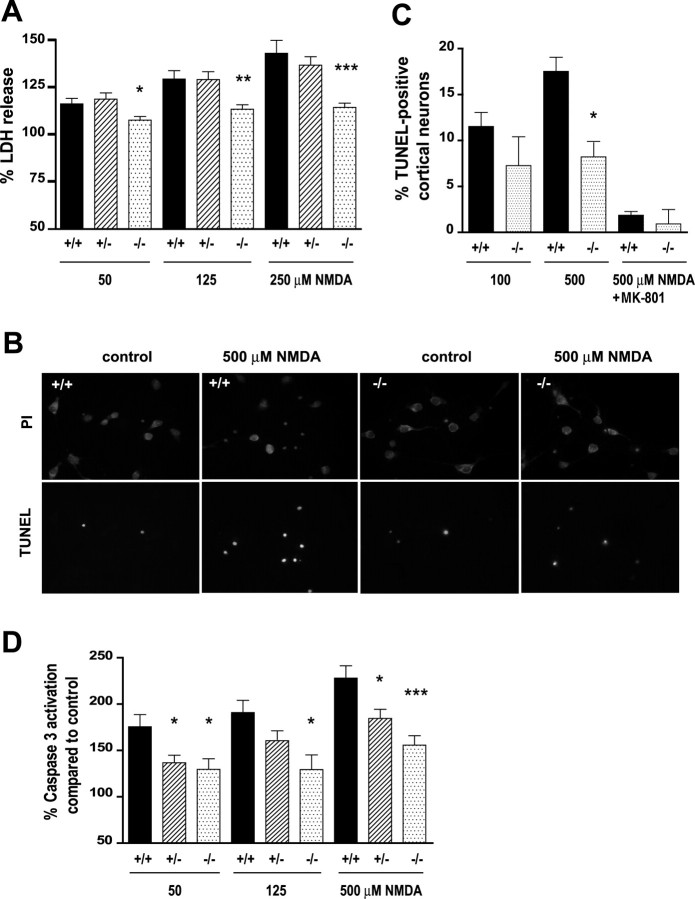
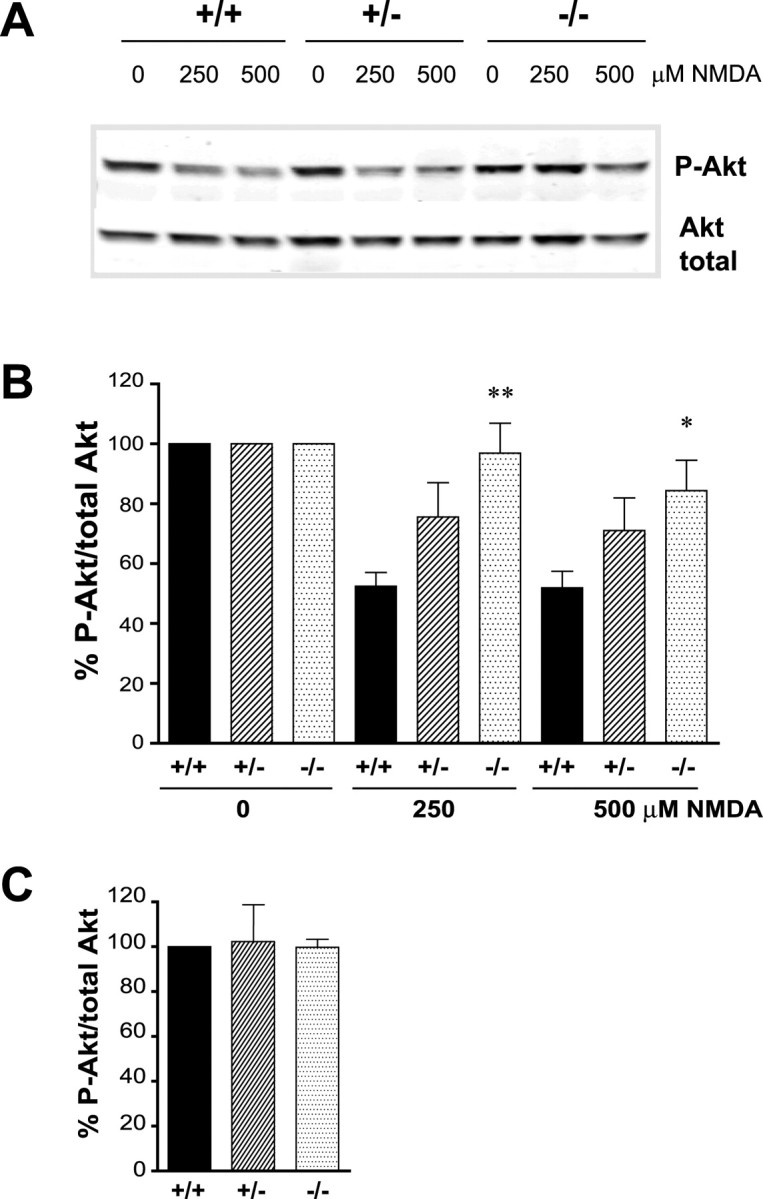
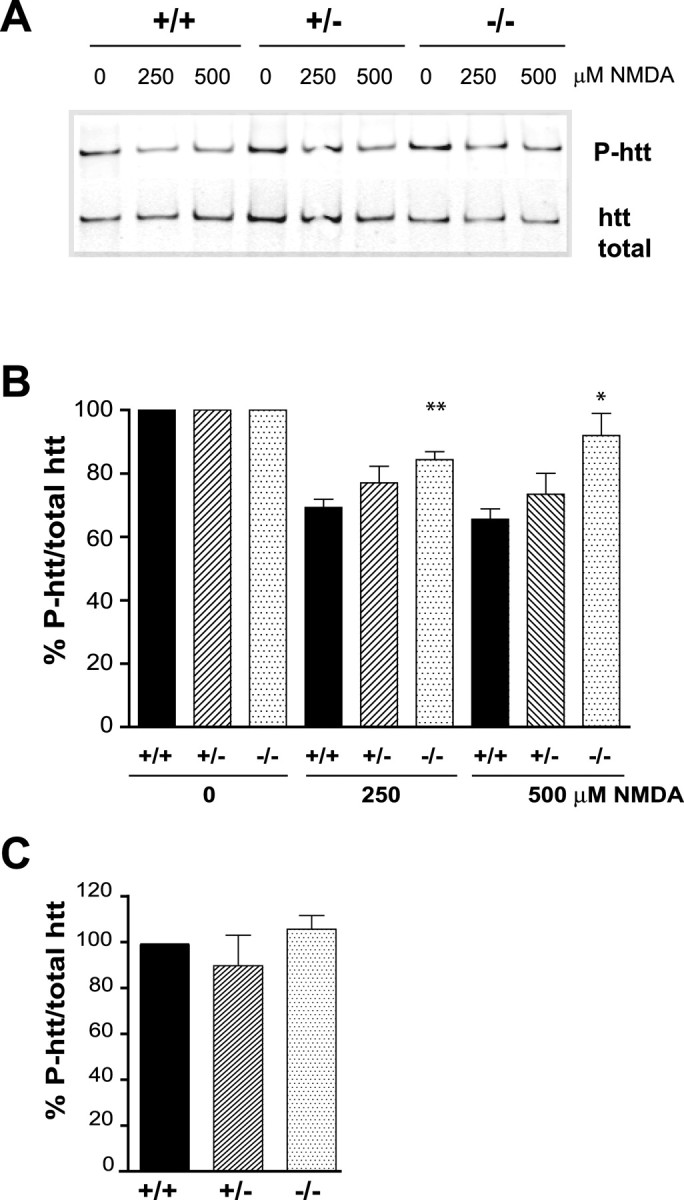
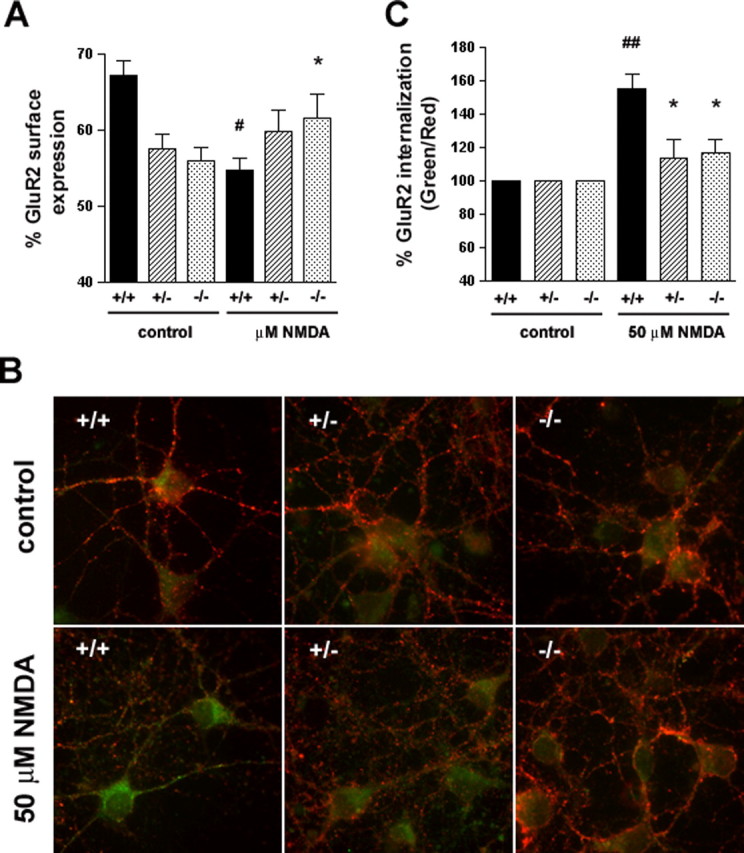
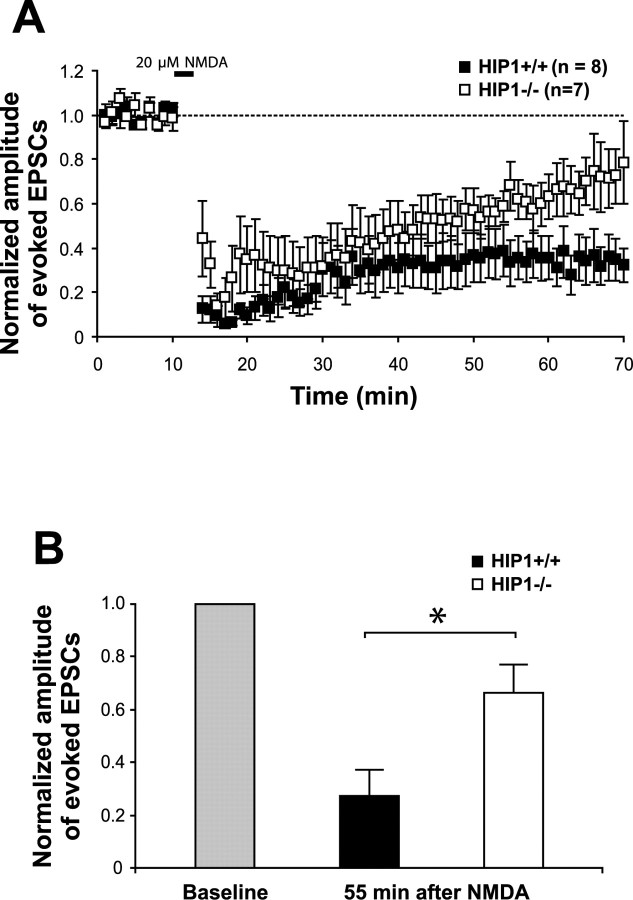
Huntingtin-interacting protein 1 (HIP1) is an endocytic adaptor protein that plays a role in clathrin-mediated endocytosis and the ligand-induced internalization of AMPA receptors (AMPARs) (Metzler et al., 2003). In the present study, we investigated the role of HIP1 in NMDA receptor (NMDAR) function by analyzing NMDA-dependent transport and NMDA-induced excitotoxicity in neurons from HIP1-/- mice. HIP1 colocalizes with NMDARs in hippocampal and cortical neurons and affinity purifies with NMDARs by GST (glutathione S-transferase) pull down and coimmunoprecipitation. A profound decrease in NMDA-induced AMPAR internalization of 75% occurs in neurons from HIP1-/- mice compared with wild type, using a quantitative single-cell-based internalization assay. This defect in NMDA-dependent removal of surface AMPARs is in agreement with the observed defect in long-term depression induction in hippocampal brain slices of HIP1-/- mice and supports a role of HIP1 in AMPAR internalization in vivo. HIP1-/- neurons are partially protected from NMDA-induced excitotoxicity as assessed by LDH (lactate dehydrogenase) release, TUNEL (terminal deoxynucleotidyl transferase-mediated biotinylated dUTP nick end labeling) and caspase-3 activation assays, which points to a role of HIP1 in NMDA-induced cell death. Interestingly, phosphorylation of Akt and its substrate huntingtin (htt) decreases during NMDA-induced excitotoxicity by 48 and 31%, respectively. This decrease is significantly modulated by HIP1, resulting in 94 and 48% changes in P-Akt and P-htt levels in HIP1-/- neurons, respectively. In summary, we have shown that HIP1 influences important NMDAR functions and that both HIP1 and htt participate in NMDA-induced cell death. These findings may provide novel insights into the cellular mechanisms underlying enhanced NMDA-induced excitotoxicity in Huntington's disease.
Figures
References
-
- Alloway PG, Howard L, Dolph PJ. The formation of stable rhodopsin-arrestin complexes induces apoptosis and photoreceptor cell degeneration. Neuron. 2000;28:129–138. - PubMed
-
- Beal MF, Kowall NW, Ellison DW, Mazurek MF, Swartz KJ, Martin JB. Replication of the neurochemical characteristics of Huntington's disease by quinolinic acid. Nature. 1986;321:168–171. - PubMed
-
- Beattie EC, Carroll RC, Yu X, Morishita W, Yasuda H, von Zastrow M, Malenka RC. Regulation of AMPA receptor endocytosis by a signaling mechanism shared with LTD. Nat Neurosci. 2000;3:1291–1300. - PubMed
-
- Carroll RC, Lissin DV, von Zastrow M, Nicoll RA, Malenka RC. Rapid redistribution of glutamate receptors contributes to long-term depression in hippocampal cultures. Nat Neurosci. 1999a;2:454–460. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials