

Anandamide regulates postnatal development of long-term synaptic plasticity in the rat dorsolateral striatum
- PMID: 17329438
- PMCID: PMC6673491
- DOI: 10.1523/JNEUROSCI.2916-06.2007
Anandamide regulates postnatal development of long-term synaptic plasticity in the rat dorsolateral striatum
Abstract
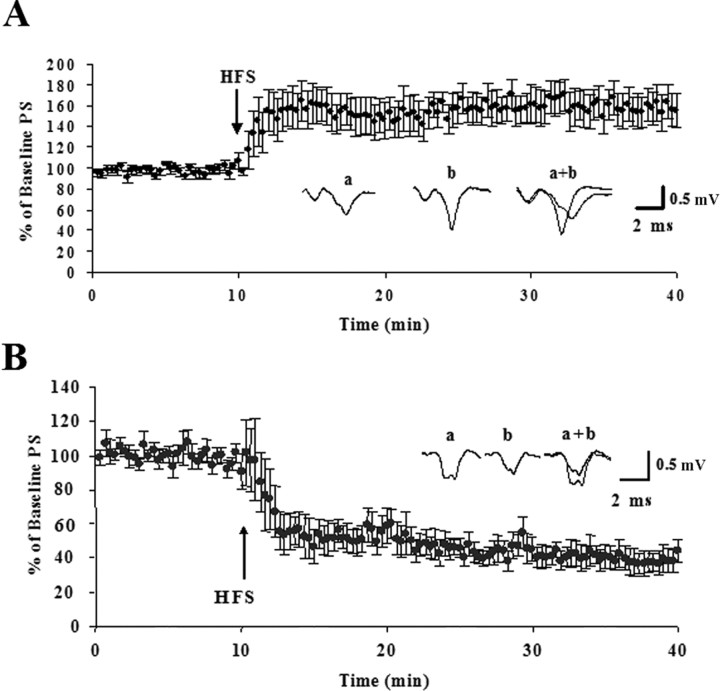
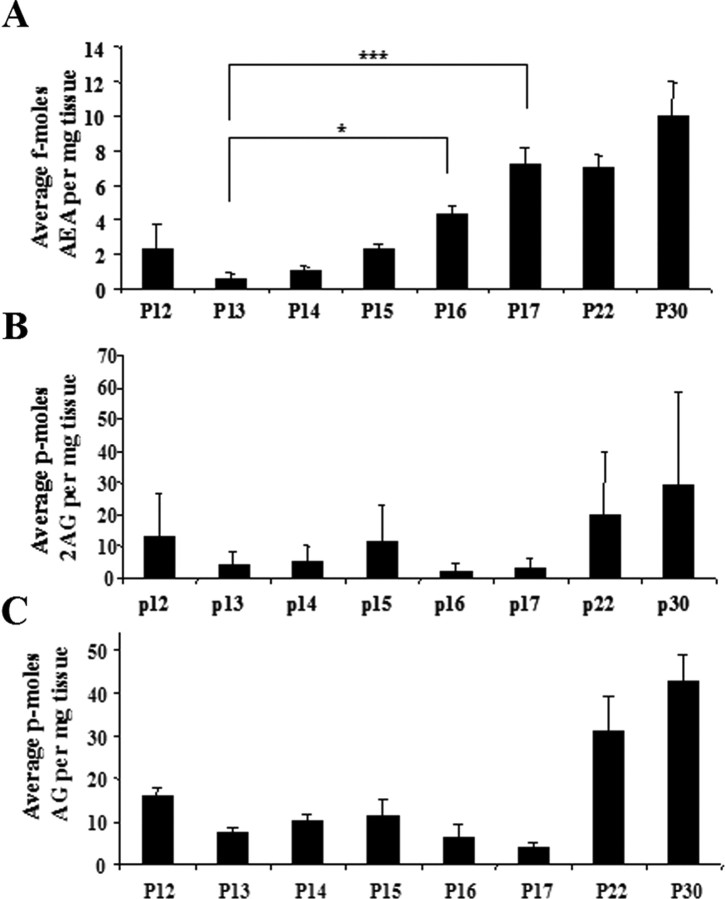
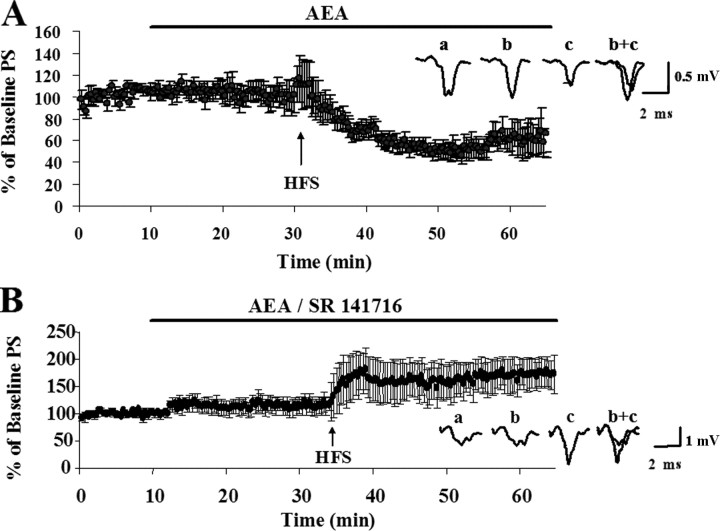
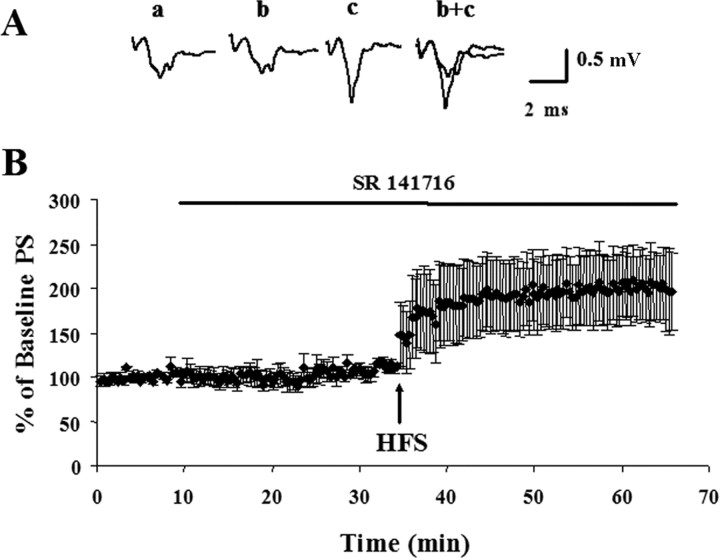
Long-term changes in synaptic efficacy produced by high-frequency stimulation (HFS) of glutamatergic afferents to the rat dorsolateral striatum exhibit heterogeneity during early stages of postnatal development. Whereas HFS most often induces striatal long-term potentiation (LTP) in rats postnatal day 12 (P12)-P14, the same stimulation tends to induce long-term depression (LTD) at ages P16-P34. Previous studies have shown that striatal LTD induction depends on retrograde endocannabinoid signaling and activation of the CB1 cannabinoid receptor. It is also known that levels of one of the primary endogenous CB1 receptor agonists, anandamide (AEA), increases during development in whole-brain samples. In the present study, we sought to determine whether this developmental increase in AEA also takes place in striatal tissue and whether increased AEA levels contribute to the postnatal switch in the response to HFS. We observed a pronounced increase in striatal levels of AEA, but not the other major endogenous cannabinoid 2-arachidonoylglycerol (2-AG), during the postnatal period characterized by the switch from LTP to LTD. Furthermore, application of synthetic AEA during HFS in field recordings of slices from P12-P14 rats allowed for induction of LTD whereas blocking the CB1 receptor during HFS in animals P16-P34 resulted in expression of LTP. However, blocking 2-AG synthesis with the DAG-lipase inhibitor tetrahydrolipstatin did not alter HFS-induced striatal LTD. In addition, synaptic depression produced by a synthetic CB1 agonist was similar across development. Together, these findings suggest that the robust developmental increase in striatal AEA may be the key factor in the emergence of HFS-induced striatal LTD.
Figures


References
-
- Ade K, Harvey-White J, Kunos G, Lovinger DM. Stimulation of dopamine D2 receptors induces the formation of anandamide (AEA) but not 2-arachidonoyl glycerol (2-AG) in rat cortical-striatal slices. Int Cannabinoid Res Soc Abstr. 2003;13:88.
-
- Barnes TD, Kubota Y, Hu D, Jin DZ, Graybiel AM. Activity of striatal neurons reflects dynamic encoding and recording of procedural memories. Nature. 2005;437:1158–1161. - PubMed
-
- Berrendero F, Sepe N, Ramos JA, Di Marzo V, Fernandez-Ruiz J. Analysis of cannabinoid receptor binding and mRNA expression and endogenous cannabinoid contents in the developing rat brain during late gestation and early postnatal period. Synapse. 1999;33:181–191. - PubMed
-
- Bisogno T, Cascia MG, Saha B, Mahadevan A, Urbani P, Minassi A, Appendino G, Saturnino C, Martin B, Razdan R, Di Marzo V. Development of the first potent and specific inhibitors of endocannabinoid biosynthesis. Biochim Biophys Acta. 2006;1761:205–212. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials