

Is the mammalian serine palmitoyltransferase a high-molecular-mass complex?
- PMID: 17331073
- PMCID: PMC1925250
- DOI: 10.1042/BJ20070025
Is the mammalian serine palmitoyltransferase a high-molecular-mass complex?
Abstract
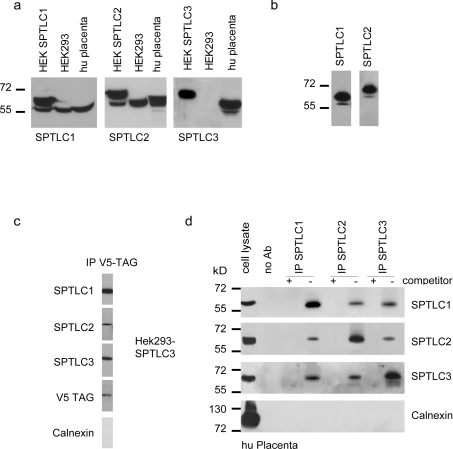
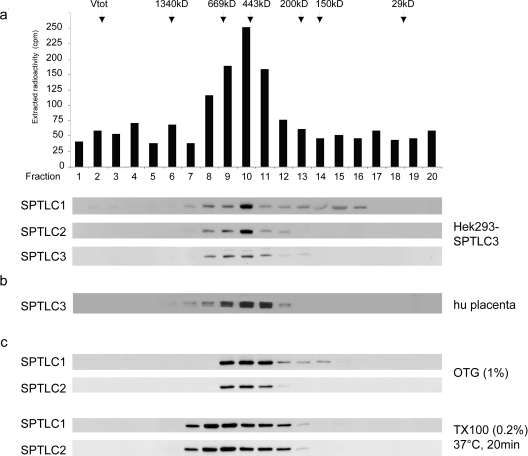
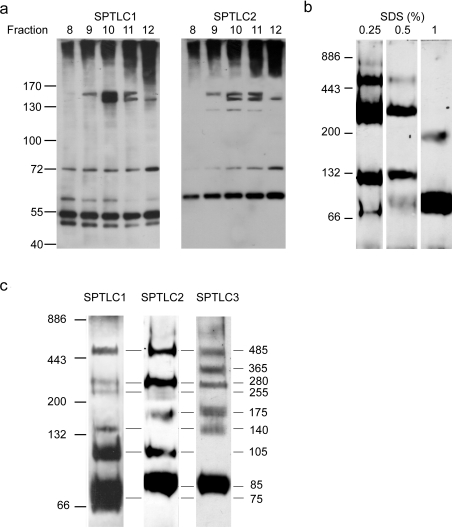
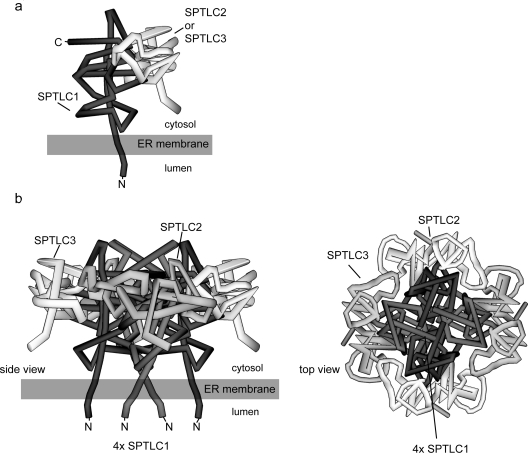
SPT (serine palmitoyltransferase) catalyses the rate-limiting step for the de novo synthesis of sphingolipids. Mammalian SPT is believed to be a heterodimer composed of two subunits, SPTLC1 and SPTLC2. We reported previously the identification of a new third SPT subunit, SPTLC3. In the present study, we have investigated the structure of the SPT complex in more detail. Pull-down assays with antibodies against SPTLC3 concomitantly co-precipitated SPTLC1 and SPTLC2 in human placenta extracts and SPTLC3 overexpressing human embryonic kidney-293 cells. By size exclusion chromatography, we determined the molecular mass of the functional SPT complex to be approx. 480 kDa. By Blue-native-PAGE experiments we demonstrated that all three SPT subunits (SPTLC1-3) are co-localized within a single SPT complex. On the basis of these results we conclude that the functional SPT is not a dimer, but a higher organized complex, composed of three distinct subunits (SPTLC1, SPTLC2 and SPTLC3) with a molecular mass of 480 kDa. The stoichiometry of SPTLC2 and SPTLC3 in this complex seems not to be fixed and is probably changed dynamically in dependence of the tissue specific SPTLC2 and SPTLC3 expression levels. Based on our own and earlier published data we propose a model of an octameric SPT structure. The observed dynamic composition of the SPT complex could provide a cellular mechanism to adjust SPT activity to tissue specific requirements in sphingolipid synthesis.
Figures
References
-
- Hannun Y. A., Luberto C., Argraves K. M. Enzymes of sphingolipid metabolism: from modular to integrative signaling. Biochemistry. 2001;40:4893–4903. - PubMed
-
- Hannun Y. A., Obeid L. M. The ceramide-centric universe of lipid-mediated cell regulation: stress encounters of the lipid kind. J. Biol. Chem. 2002;277:25847–25850. - PubMed
-
- Merrill A. H., Jr, Nixon D. W., Williams R. D. Activities of serine palmitoyltransferase (3-ketosphinganine synthase) in microsomes from different rat tissues. J. Lipid Res. 1985;26:617–622. - PubMed
-
- Hojjati M. R., Li Z., Jiang X. C. Serine palmitoyl-CoA transferase (SPT) deficiency and sphingolipid levels in mice. Biochim. Biophys. Acta. 2005;1737:44–51. - PubMed
-
- Hanada K. Serine palmitoyltransferase, a key enzyme of sphingolipid metabolism. Biochim. Biophys. Acta. 2003;1632:16–30. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases