

An improved definition of the RNA-binding specificity of SECIS-binding protein 2, an essential component of the selenocysteine incorporation machinery
- PMID: 17332014
- PMCID: PMC1874613
- DOI: 10.1093/nar/gkm066
An improved definition of the RNA-binding specificity of SECIS-binding protein 2, an essential component of the selenocysteine incorporation machinery
Abstract
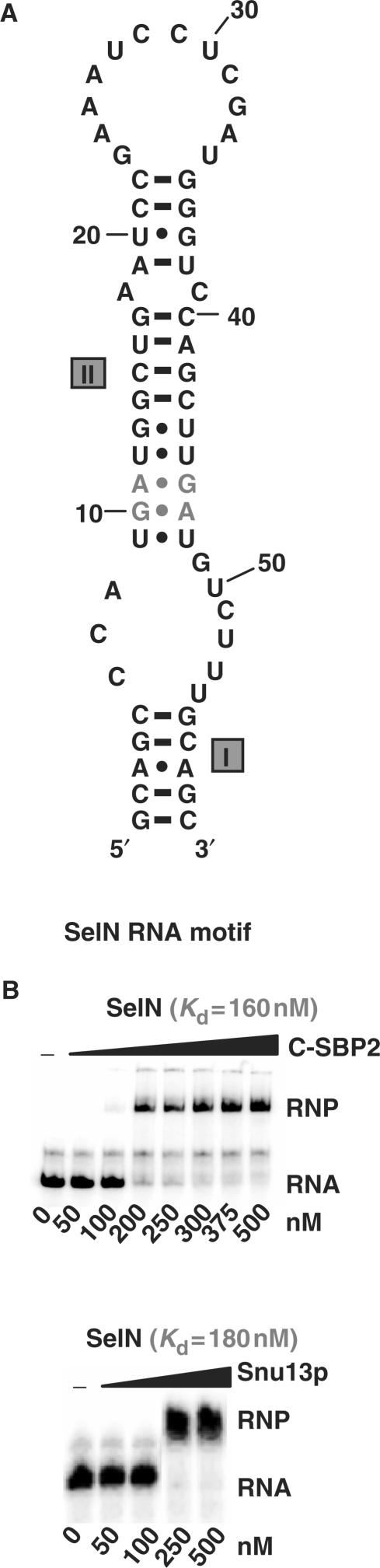
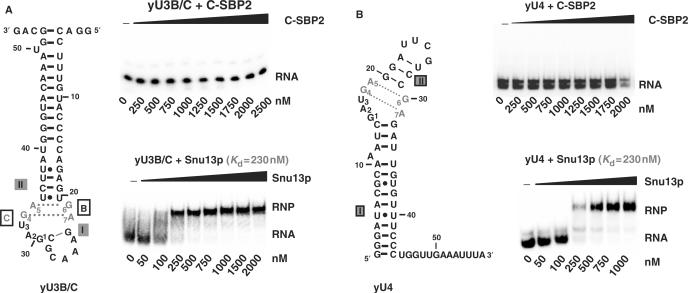
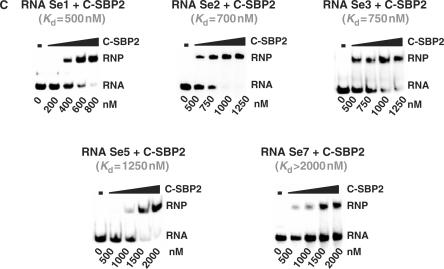
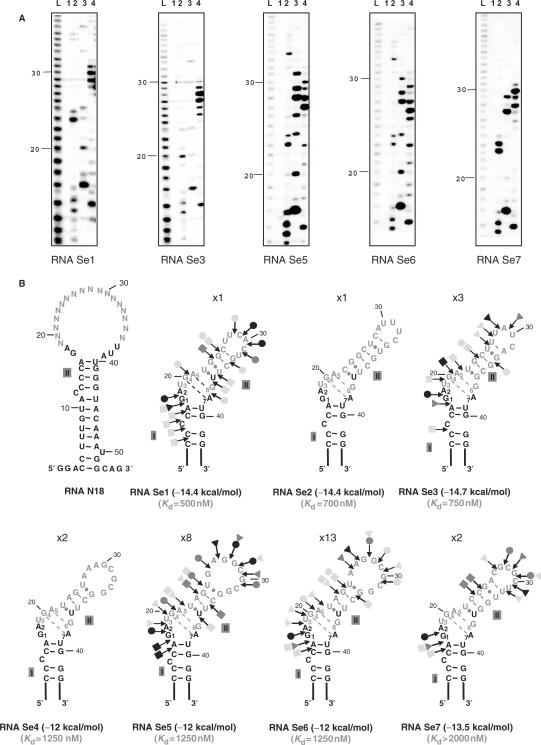
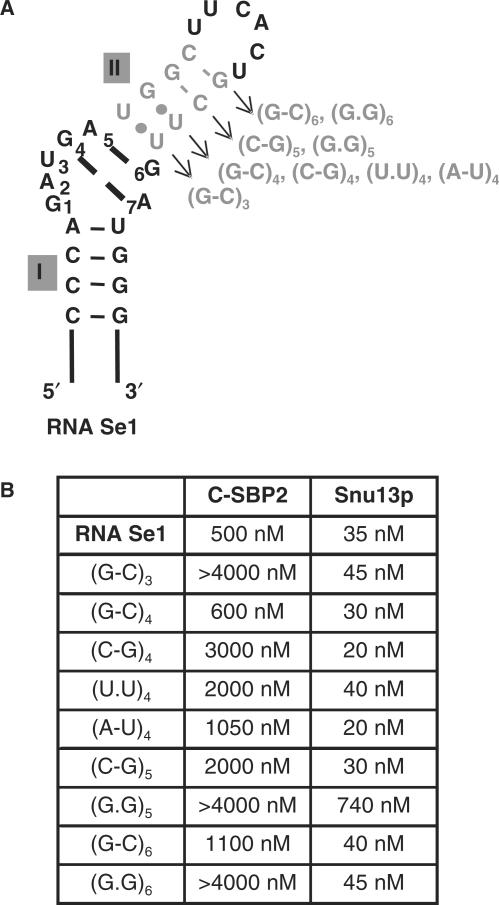
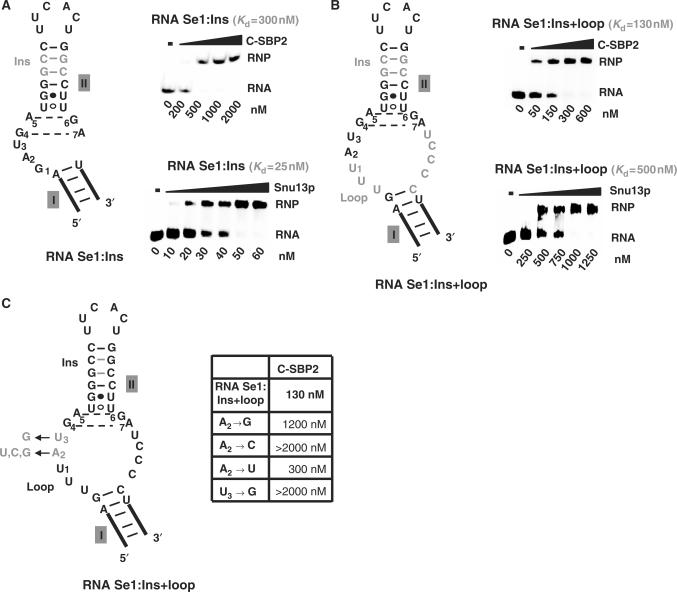
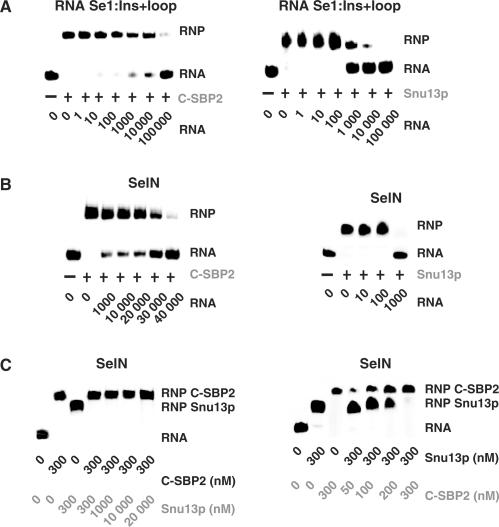
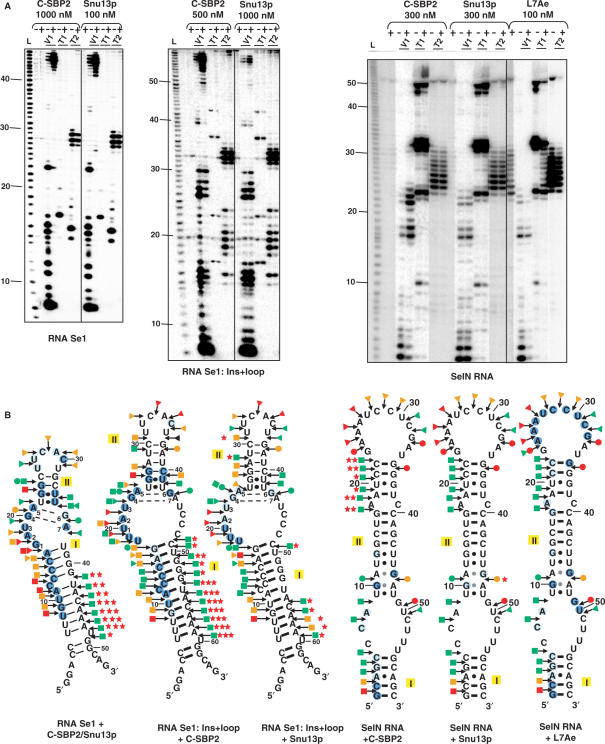
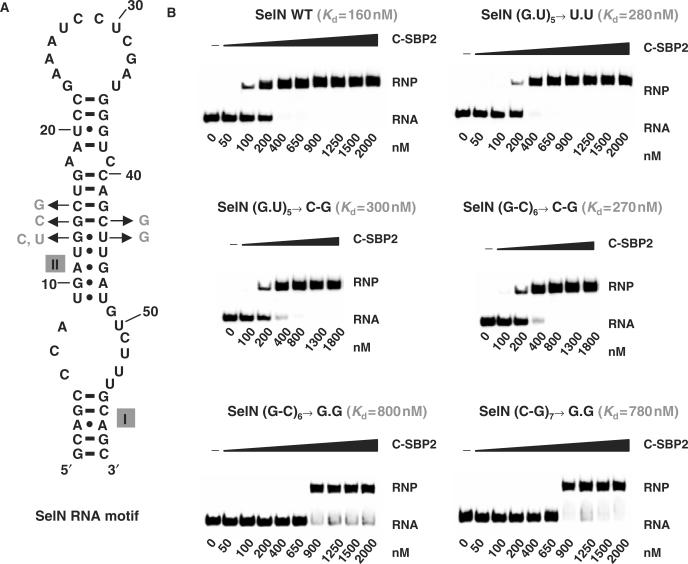
By binding to SECIS elements located in the 3'-UTR of selenoprotein mRNAs, the protein SBP2 plays a key role in the assembly of the selenocysteine incorporation machinery. SBP2 contains an L7Ae/L30 RNA-binding domain similar to that of protein 15.5K/Snu13p, which binds K-turn motifs with a 3-nt bulge loop closed by a tandem of G.A and A.G pairs. Here, by SELEX experiments, we demonstrate the capacity of SBP2 to bind such K-turn motifs with a protruding U residue. However, we show that conversion of the bulge loop into an internal loop reinforces SBP2 affinity and to a greater extent RNP stability. Opposite variations were found for Snu13p. Accordingly, footprinting assays revealed strong contacts of SBP2 with helices I and II and the 5'-strand of the internal loop, as opposed to the loose interaction of Snu13p. Our data also identifies new determinants for SBP2 binding which are located in helix II. Among the L7Ae/L30 family members, these determinants are unique to SBP2. Finally, in accordance with functional data on SECIS elements, the identity of residues at positions 2 and 3 in the loop influences SBP2 affinity. Altogether, the data provide a very precise definition of the SBP2 RNA specificity.
Figures

References
-
- Wimberly BT, Brodersen DE, Clemons WM, Jr, Morgan-Warren RJ, Carter AP, Vonrhein C, Hartsch T, Ramakrishnan V. Structure of the 30S ribosomal subunit. Nature. 2000;407:327–339. - PubMed
-
- Schluenzen F, Tocilj A, Zarivach R, Harms J, Gluehmann M, Janell D, Bashan A, Bartels H, Agmon I, et al. Structure of functionally activated small ribosomal subunit at 3.3 angstroms resolution. Cell. 2000;102:615–623. - PubMed
-
- Ban N, Nissen P, Hansen J, Moore PB, Steitz TA. The complete atomic structure of the large ribosomal subunit at 2.4 Å resolution. Science. 2000;289:905–920. - PubMed
-
- Vidovic I, Nottrott S, Hartmuth K, Luhrmann R, Ficner R. Crystal structure of the spliceosomal 15.5kD protein bound to a U4 snRNA fragment. Mol. Cell. 2000;6:1331–1342. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
