

Alternative splicing and bioinformatic analysis of human U12-type introns
- PMID: 17332017
- PMCID: PMC1874599
- DOI: 10.1093/nar/gkm026
Alternative splicing and bioinformatic analysis of human U12-type introns
Abstract
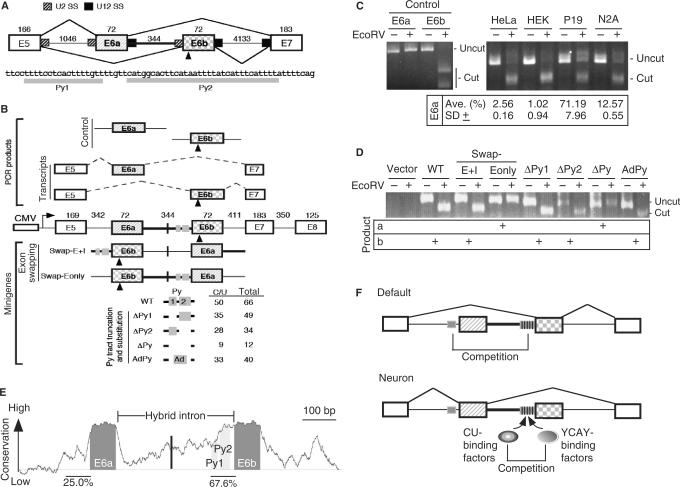
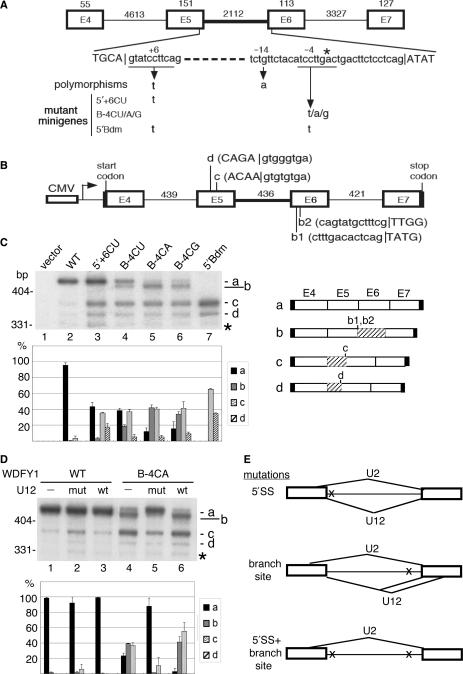
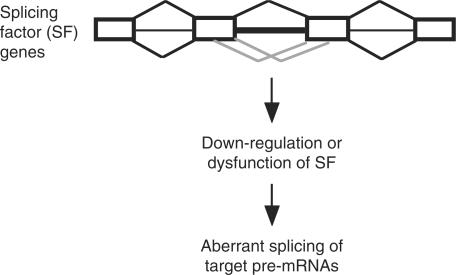
U12-type introns exist, albeit rarely, in a variety of multicellular organisms. Splicing of U12 intron-containing precursor mRNAs takes place in the U12-type spliceosome that is distinct from the major U2-type spliceosome. Due to incompatibility of these two spliceosomes, alternative splicing involving a U12-type intron may give rise to a relatively complicated impact on gene expression. We studied alternative U12-type intron splicing in an attempt to gain more mechanistic insights. First, we characterized mutually exclusive exon selection of the human JNK2 gene, which involves an unusual intron possessing the U12-type 5' splice site and the U2-type 3' splice site. We demonstrated that the long and evolutionary conserved polypyrimidine tract of this hybrid intron provides important signals for inclusion of its downstream alternative exon. In addition, we examined the effects of single nucleotide polymorphisms in the human WDFY1 U12-type intron on pre-mRNA splicing. These results provide mechanistic implications on splice-site selection of U12-type intron splicing. We finally discuss the potential effects of splicing of a U12-type intron with genetic defects or within a set of genes encoding RNA processing factors on global gene expression.
Figures
Similar articles
-
RS domain-splicing signal interactions in splicing of U12-type and U2-type introns.Nat Struct Mol Biol. 2007 Jul;14(7):597-603. doi: 10.1038/nsmb1263. Epub 2007 Jul 1. Nat Struct Mol Biol. 2007. PMID: 17603499
-
An LKB1 AT-AC intron mutation causes Peutz-Jeghers syndrome via splicing at noncanonical cryptic splice sites.Nat Struct Mol Biol. 2005 Jan;12(1):54-9. doi: 10.1038/nsmb873. Epub 2004 Dec 19. Nat Struct Mol Biol. 2005. PMID: 15608654
-
Human GC-AG alternative intron isoforms with weak donor sites show enhanced consensus at acceptor exon positions.Nucleic Acids Res. 2001 Jun 15;29(12):2581-93. doi: 10.1093/nar/29.12.2581. Nucleic Acids Res. 2001. PMID: 11410667 Free PMC article.
-
Splicing of a rare class of introns by the U12-dependent spliceosome.Biol Chem. 2005 Aug;386(8):713-24. doi: 10.1515/BC.2005.084. Biol Chem. 2005. PMID: 16201866 Review.
-
Alternative splicing of U12-type introns.Front Biosci. 2008 Jan 1;13:1681-90. doi: 10.2741/2791. Front Biosci. 2008. PMID: 17981659 Review.
Cited by
-
Chromosomal instability by mutations in the novel minor spliceosome component CENATAC.EMBO J. 2021 Jul 15;40(14):e106536. doi: 10.15252/embj.2020106536. Epub 2021 May 19. EMBO J. 2021. PMID: 34009673 Free PMC article.
-
New insights into minor splicing-a transcriptomic analysis of cells derived from TALS patients.RNA. 2019 Sep;25(9):1130-1149. doi: 10.1261/rna.071423.119. Epub 2019 Jun 7. RNA. 2019. PMID: 31175170 Free PMC article.
-
The Arabidopsis U12-type spliceosomal protein U11/U12-31K is involved in U12 intron splicing via RNA chaperone activity and affects plant development.Plant Cell. 2010 Dec;22(12):3951-62. doi: 10.1105/tpc.110.079103. Epub 2010 Dec 10. Plant Cell. 2010. PMID: 21148817 Free PMC article.
-
Convergent evolution of twintron-like configurations: One is never enough.RNA Biol. 2015;12(12):1275-88. doi: 10.1080/15476286.2015.1103427. RNA Biol. 2015. PMID: 26513606 Free PMC article. Review.
-
Spliceosomal gene mutations in myelodysplasia: molecular links to clonal abnormalities of hematopoiesis.Genes Dev. 2016 May 1;30(9):989-1001. doi: 10.1101/gad.278424.116. Genes Dev. 2016. PMID: 27151974 Free PMC article. Review.
References
-
- Tarn WY, Steitz JA. Pre-mRNA splicing: the discovery of a new spliceosome doubles the challenge. Trends Biochem. Sci. 1997;22:132–137. - PubMed
-
- Burge CB, Tuschl T, Sharp PA. Splicing of precursor to mRNA by the spliceosomes. In: Gesteland RF, Cech T, Aktins JF, editors. The RNA World II. Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press; 1999.
-
- Sharp PA, Burge CB. Classification of intron: U2-type or U12-type. Cell. 1997;91:875–879. - PubMed
-
- Will CL, Luhrmann R. Splicing of a rare class of intron by the U12-dependent spliceosome. J. Biol. Chem. 2005;386:713–724. - PubMed
-
- Patel AA, Steitz JA. Splicing double: insights from the second spliceosome. Nat. Rev. Mol. Cell. Biol. 2003;4:960–970. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
