

Stationary phase reorganisation of the Escherichia coli transcription machinery by Crl protein, a fine-tuner of sigmas activity and levels
- PMID: 17332743
- PMCID: PMC1829388
- DOI: 10.1038/sj.emboj.7601629
Stationary phase reorganisation of the Escherichia coli transcription machinery by Crl protein, a fine-tuner of sigmas activity and levels
Abstract
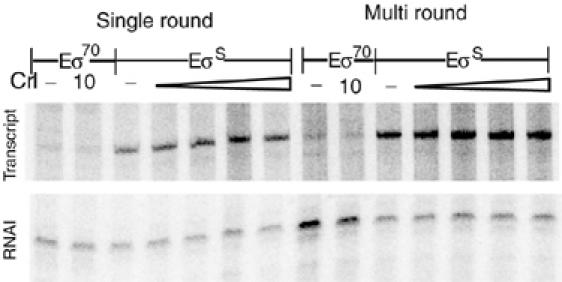
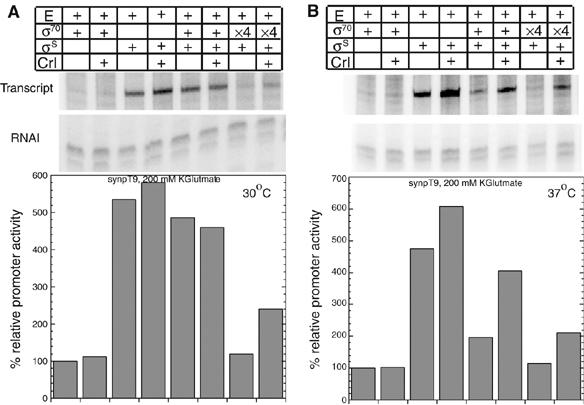
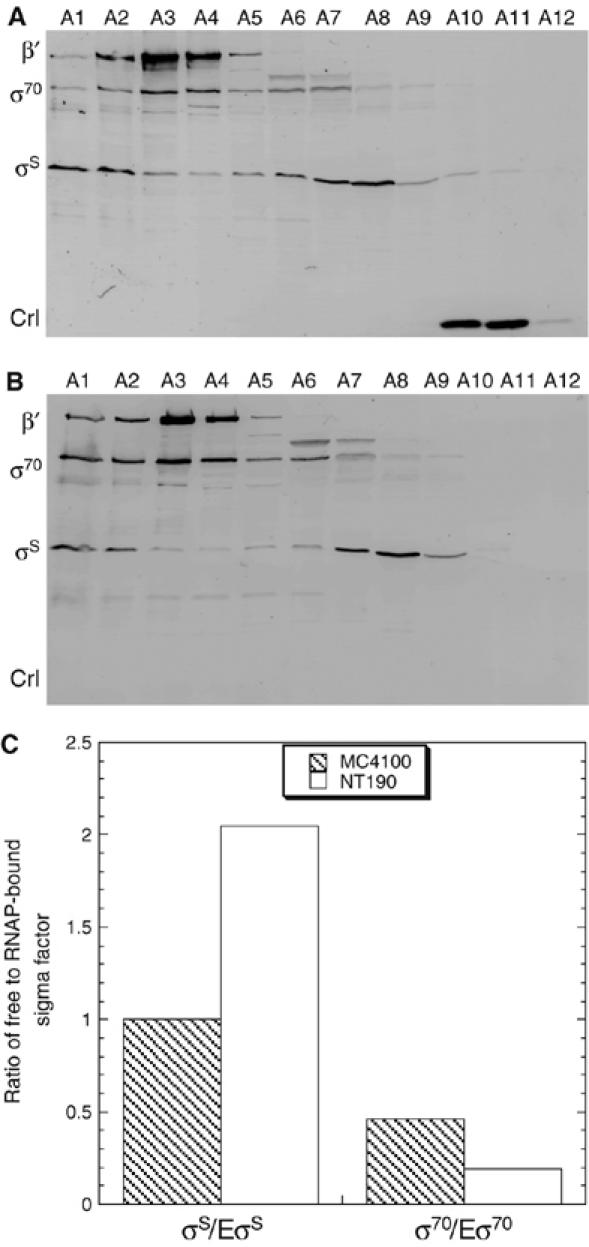
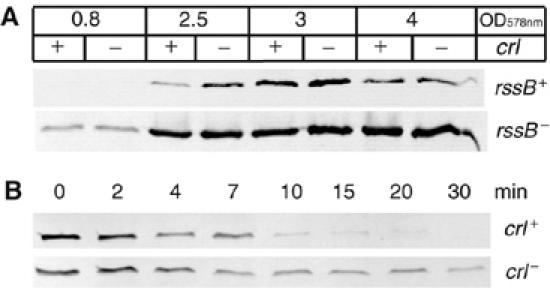
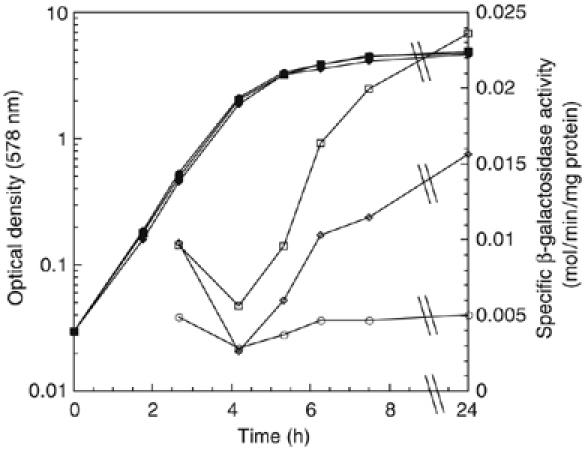
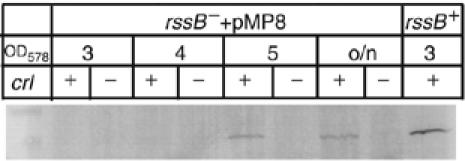
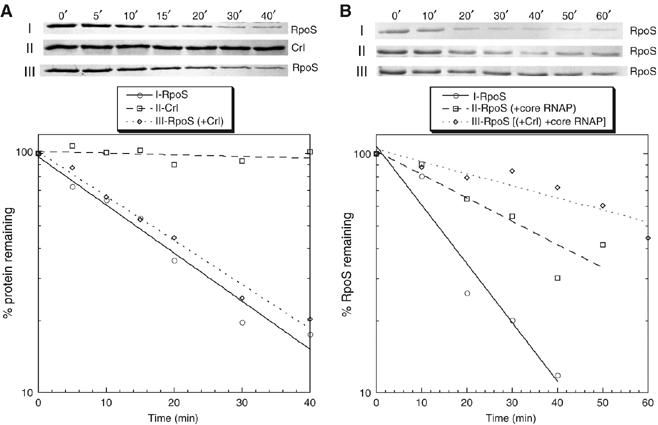
Upon environmental changes, bacteria reschedule gene expression by directing alternative sigma factors to core RNA polymerase (RNAP). This sigma factor switch is achieved by regulating relative amounts of alternative sigmas and by decreasing the competitiveness of the dominant housekeeping sigma(70). Here we report that during stationary phase, the unorthodox Crl regulator supports a specific sigma factor, sigma(S) (RpoS), in its competition with sigma(70) for core RNAP by increasing the formation of sigma(S)-containing RNAP holoenzyme, Esigma(S). Consistently, Crl has a global regulatory effect in stationary phase gene expression exclusively through sigma(S), that is, on sigma(S)-dependent genes only. Not a specific promoter motif, but sigma(S) availability determines the ability of Crl to exert its function, rendering it of major importance at low sigma(S) levels. By promoting the formation of Esigma(S), Crl also affects partitioning of sigma(S) between RNAP core and the proteolytic sigma(S)-targeting factor RssB, thereby playing a dual role in fine-tuning sigma(S) proteolysis. In conclusion, Crl has a key role in reorganising the Escherichia coli transcriptional machinery and global gene expression during entry into stationary phase.
Figures
Similar articles
-
The P1 promoter of the Escherichia coli rpoH gene is utilized by sigma 70 -RNAP or sigma s -RNAP depending on growth phase.FEMS Microbiol Lett. 2009 Feb;291(1):65-72. doi: 10.1111/j.1574-6968.2008.01436.x. Epub 2008 Dec 3. FEMS Microbiol Lett. 2009. PMID: 19076234
-
The response regulator RssB controls stability of the sigma(S) subunit of RNA polymerase in Escherichia coli.EMBO J. 1996 Mar 15;15(6):1333-9. EMBO J. 1996. PMID: 8635466 Free PMC article.
-
The molecular basis of selective promoter activation by the sigmaS subunit of RNA polymerase.Mol Microbiol. 2007 Mar;63(5):1296-306. doi: 10.1111/j.1365-2958.2007.05601.x. Mol Microbiol. 2007. PMID: 17302812 Review.
-
Sigma and RNA polymerase: an on-again, off-again relationship?Mol Cell. 2005 Nov 11;20(3):335-45. doi: 10.1016/j.molcel.2005.10.015. Mol Cell. 2005. PMID: 16285916 Review.
-
sigma factor selectivity of Escherichia coli RNA polymerase: role for CRP, IHF and lrp transcription factors.EMBO J. 2000 Jun 15;19(12):3028-37. doi: 10.1093/emboj/19.12.3028. EMBO J. 2000. PMID: 10856246 Free PMC article.
Cited by
-
The actinobacterial transcription factor RbpA binds to the principal sigma subunit of RNA polymerase.Nucleic Acids Res. 2013 Jun;41(11):5679-91. doi: 10.1093/nar/gkt277. Epub 2013 Apr 19. Nucleic Acids Res. 2013. PMID: 23605043 Free PMC article.
-
Activation of σ20-dependent recombination and horizontal gene transfer in Mycoplasma genitalium.DNA Res. 2018 Aug 1;25(4):383-393. doi: 10.1093/dnares/dsy011. DNA Res. 2018. PMID: 29659762 Free PMC article.
-
Co-ordinated regulation of the extracytoplasmic stress factor, sigmaE, with other Escherichia coli sigma factors by (p)ppGpp and DksA may be achieved by specific regulation of individual holoenzymes.Mol Microbiol. 2014 Aug;93(3):479-93. doi: 10.1111/mmi.12674. Epub 2014 Jul 10. Mol Microbiol. 2014. PMID: 24946009 Free PMC article.
-
Mycobacterium RbpA cooperates with the stress-response σB subunit of RNA polymerase in promoter DNA unwinding.Nucleic Acids Res. 2014;42(16):10399-408. doi: 10.1093/nar/gku742. Epub 2014 Aug 13. Nucleic Acids Res. 2014. PMID: 25122744 Free PMC article.
-
Survival guide: Escherichia coli in the stationary phase.Acta Naturae. 2015 Oct-Dec;7(4):22-33. Acta Naturae. 2015. PMID: 26798489 Free PMC article.
References
-
- Arnqvist A, Olsen A, Pfeifer J, Russell DG, Normark S (1992) The Crl protein activates cryptic genes for curli formation and fibronectin binding in Escherichia coli HB101. Mol Microbiol 6: 2443–2452 - PubMed
-
- Becker G, Hengge-Aronis R (2001) What makes an Escherichia coli promoter σS-dependent? Role of the −13/−14 nucleotide promoter positions and region 2.5 of σS. Mol Microbiol 39: 1153–1165 - PubMed
-
- Becker G, Klauck E, Hengge-Aronis R (2000) The response regulator RssB, a recognition factor for sigmas proteolysis in Escherichia coli, can act like an anti-sigmas factor. Mol Microbiol 35: 657–666 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
