

Notch3 and the Notch3-upregulated RNA-binding protein HuD regulate Ikaros alternative splicing
- PMID: 17332745
- PMCID: PMC1829386
- DOI: 10.1038/sj.emboj.7601626
Notch3 and the Notch3-upregulated RNA-binding protein HuD regulate Ikaros alternative splicing
Abstract
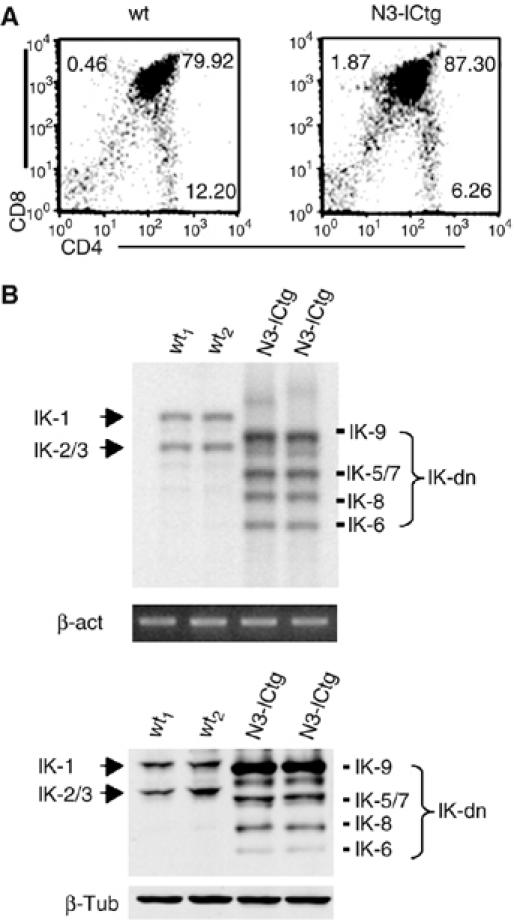
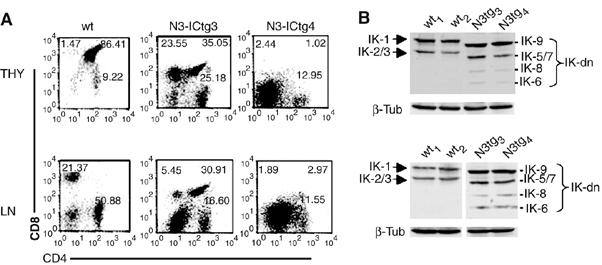
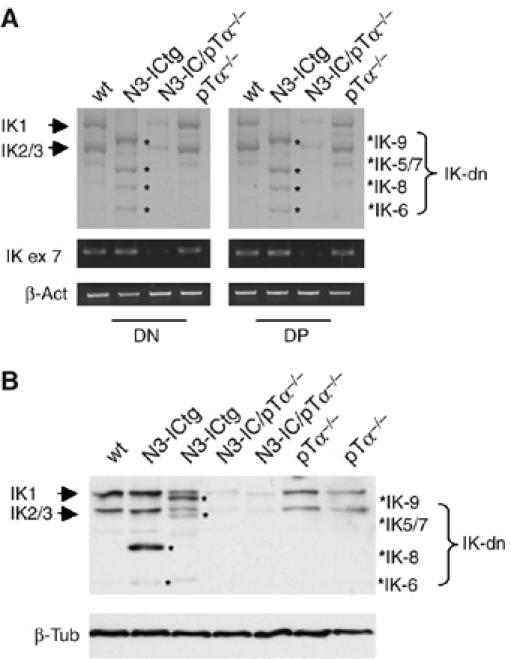
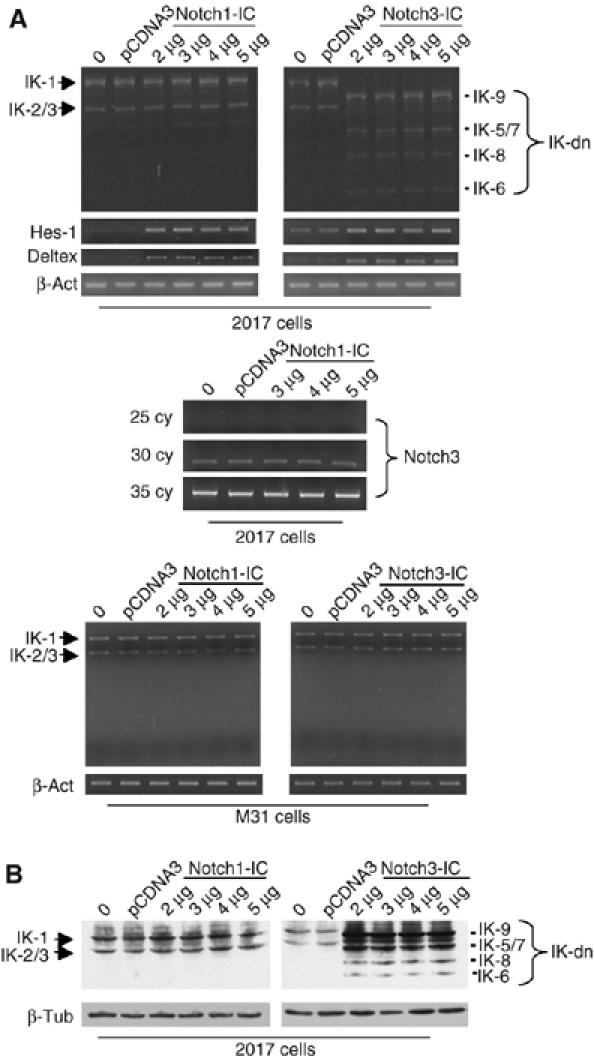
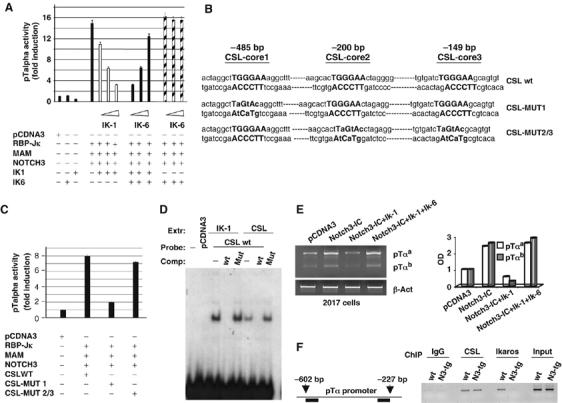
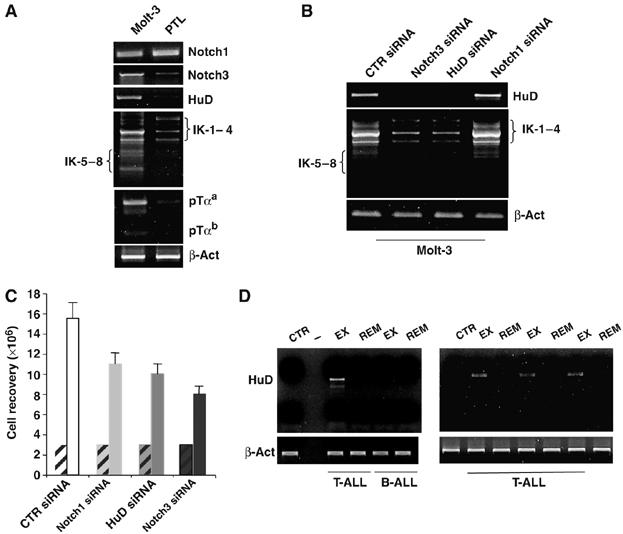
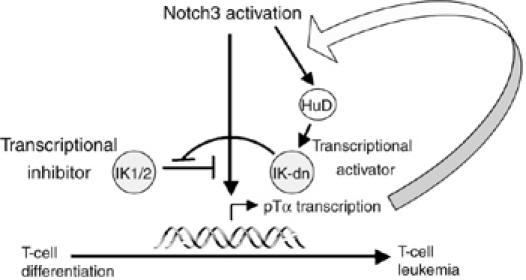
Constitutive activation of the transmembrane receptor, Notch3, and loss of function of the hematopoietic transcription repressor, Ikaros (IK), play direct roles in T-cell differentiation and leukemogenesis that are dependent on pre-T-cell receptor (pre-TCR) signaling. We demonstrate the occurrence of crosstalk between Notch3 and IK that results in transcriptional regulation of the gene encoding the pTalpha chain of the pre-TCR. We also show that, in the presence of the pre-TCR, constitutive activation of Notch3 in thymocytes causes increased expression of dominantnegative non-DNA-binding IK isoforms, which are able to restrain the IK inhibition of Notch3's transcriptional activation of pTalpha. This effect appears to be mediated by Notch3's pre-TCR-dependent upregulation of the RNA-binding protein, HuD. Notch3 signaling thus appears to play a critical role in the diminished IK activity described in several lymphoid leukemias. By exerting transcription-activating and transcription-repressing effects on the pTalpha promoter, Notch3 and IK cooperate in the fine-tuning of pre-TCR expression and function, which has important implications for the regulation of thymocyte differentiation and proliferation.
Figures

References
-
- Bellavia D, Campese AF, Alesse E, Vacca A, Felli MP, Balestri A, Stoppacciaro A, Tiveron C, Tatangelo L, Giovarelli M, Gaetano C, Ruco L, Hoffman ES, Hayday AC, Lendahl U, Frati L, Gulino A, Screpanti I (2000) Constitutive activation of NF-kappaB and T-cell leukemia/lymphoma in Notch3 transgenic mice. EMBO J 19: 3337–3348 - PMC - PubMed
-
- Bellavia D, Campese AF, Checquolo S, Balestri A, Biondi A, Cazzaniga G, Lendahl U, Fehling HJ, Hayday AC, Frati L, von Boehmer H, Gulino A, Screpanti I (2002) Combined expression of pTalpha and Notch3 in T cell leukemia identifies the requirement of preTCR for leukemogenesis. Proc Natl Acad Sci USA 99: 3788–3793 - PMC - PubMed
-
- Beverly LJ, Capobianco AJ (2003) Perturbation of Ikaros isoform selection by MLV integration is a cooperative event in Notch(IC)-induced T cell leukemogenesis. Cancer Cell 3: 551–564 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
