

Analysis of interactions in a tapasin/class I complex provides a mechanism for peptide selection
- PMID: 17332746
- PMCID: PMC1829385
- DOI: 10.1038/sj.emboj.7601624
Analysis of interactions in a tapasin/class I complex provides a mechanism for peptide selection
Abstract
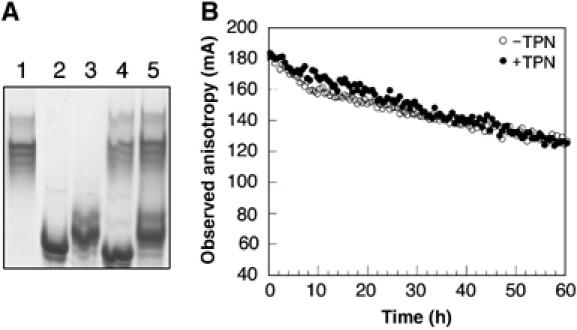
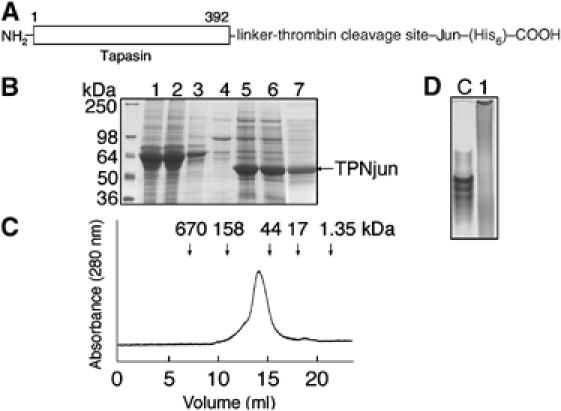
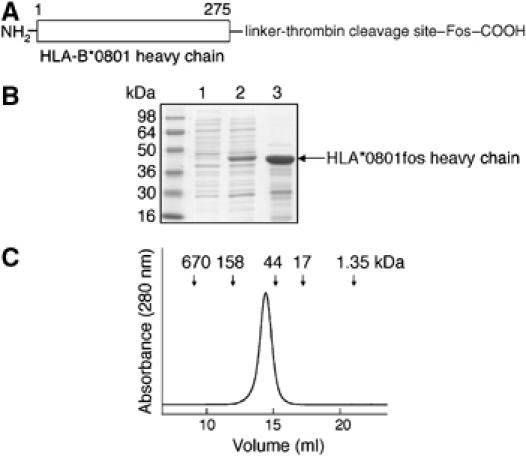
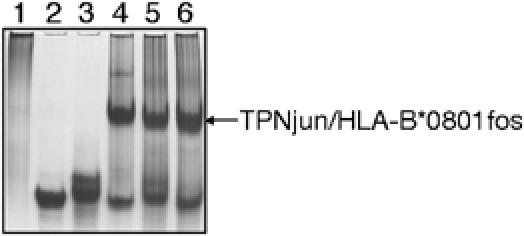
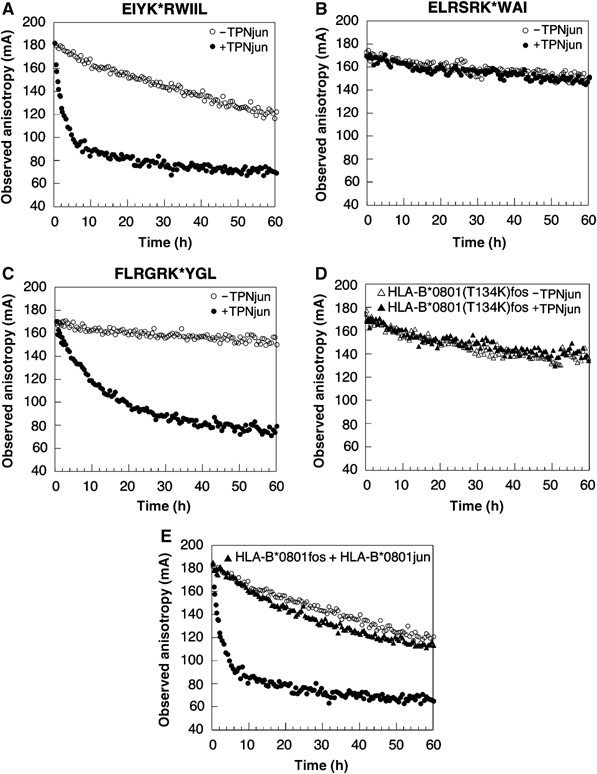
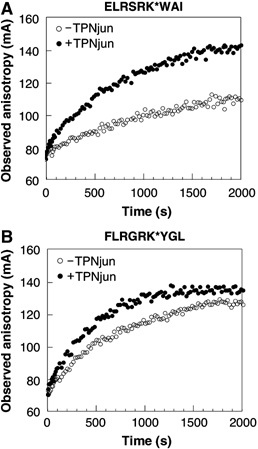
We examined interactions in a soluble tapasin (TPN)/HLA-B*0801 complex to gain mechanistic insights into the functions of TPN. Results show that TPN acts as a chaperone by increasing the ratio of active-to-inactive peptide-deficient HLA-B*0801 molecules in solution. TPN causes peptides to associate and dissociate faster owing to its effect on widening the binding groove of HLA-B*0801 molecules. Our data indicate that a TPN-assisted mechanism of peptide selection relies on disruption of conserved hydrogen bonds at the C-terminal end of the groove. Peptide sequence-dependent interactions along the entire length of the groove also play a role in this mechanism. We suggest that TPN influences presentation of antigenic peptides according to a mechanistically complicated process in which bound candidate peptides that are unable to conformationally disengage TPN from class I molecules are excluded from the repertoire. Overall, these studies unify our understanding of the functions of TPN.
Figures
References
-
- Barber LD, Howarth M, Bowness P, Elliott T (2001) The quantity of naturally processed peptides stably bound by HLA-A*0201 is significantly reduced in the absence of tapasin. Tissue Antigens 58: 363–468 - PubMed
-
- Barnden MJ, Purcell AW, Gorman JJ, McCluskey J (2000) Tapasin-mediated retention and optimization of peptide ligands during the assembly of class I molecules. J Immunol 165: 322–330 - PubMed
-
- Belmares MP, Busch R, Wucherpfennig KW, McConnell HM, Mellins ED (2002) Structural factors contributing to DM susceptibility of MHC class II/peptide complexes. J Immunol 169: 5109–5117 - PubMed
-
- Binz AK, Rodriguez RC, Biddison WE, Baker BM (2003) Thermodynamic and kinetic analysis of a peptide-class I MHC interaction highlights the noncovalent nature and conformational dynamics of the class I heterotrimer. Biochemistry 42: 4954–4961 - PubMed
-
- Bouvier M, Wiley DC (1994) Importance of peptide amino and carboxyl termini to the stability of MHC class I molecules. Science 265: 398–402 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
