

H3 histamine receptor agonist inhibits biliary growth of BDL rats by downregulation of the cAMP-dependent PKA/ERK1/2/ELK-1 pathway
- PMID: 17334413
- PMCID: PMC3751000
- DOI: 10.1038/labinvest.3700533
H3 histamine receptor agonist inhibits biliary growth of BDL rats by downregulation of the cAMP-dependent PKA/ERK1/2/ELK-1 pathway
Abstract
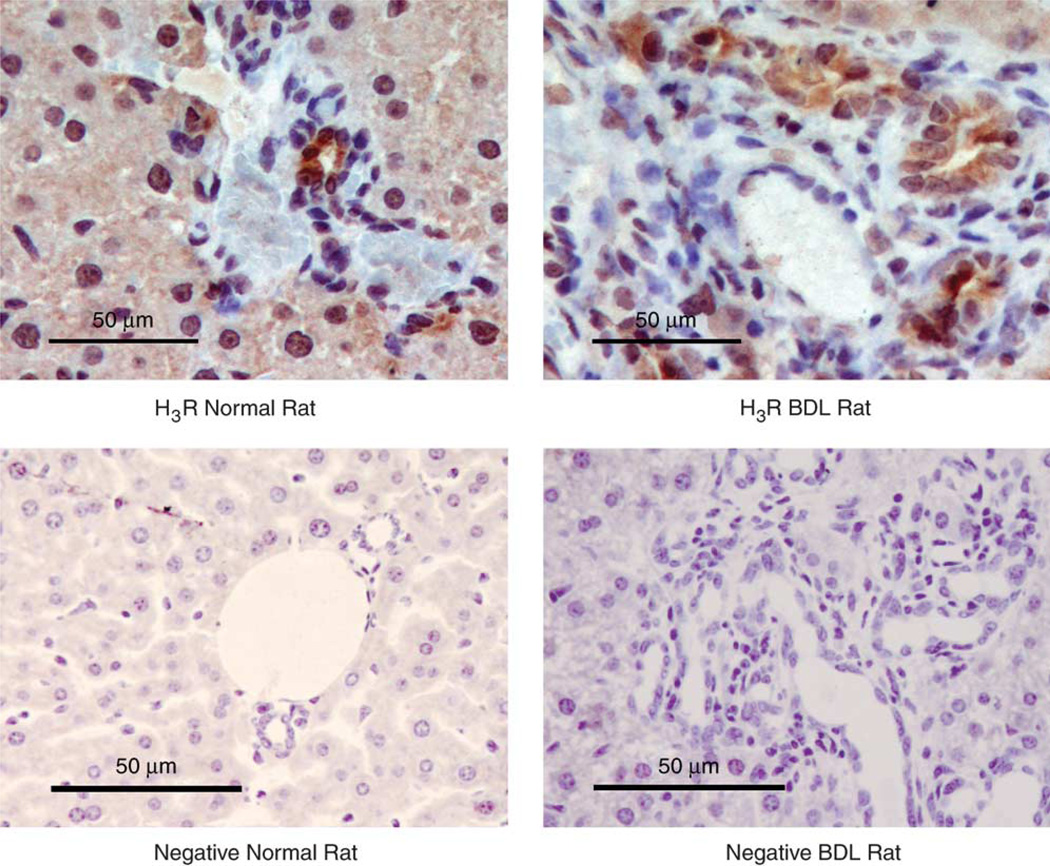
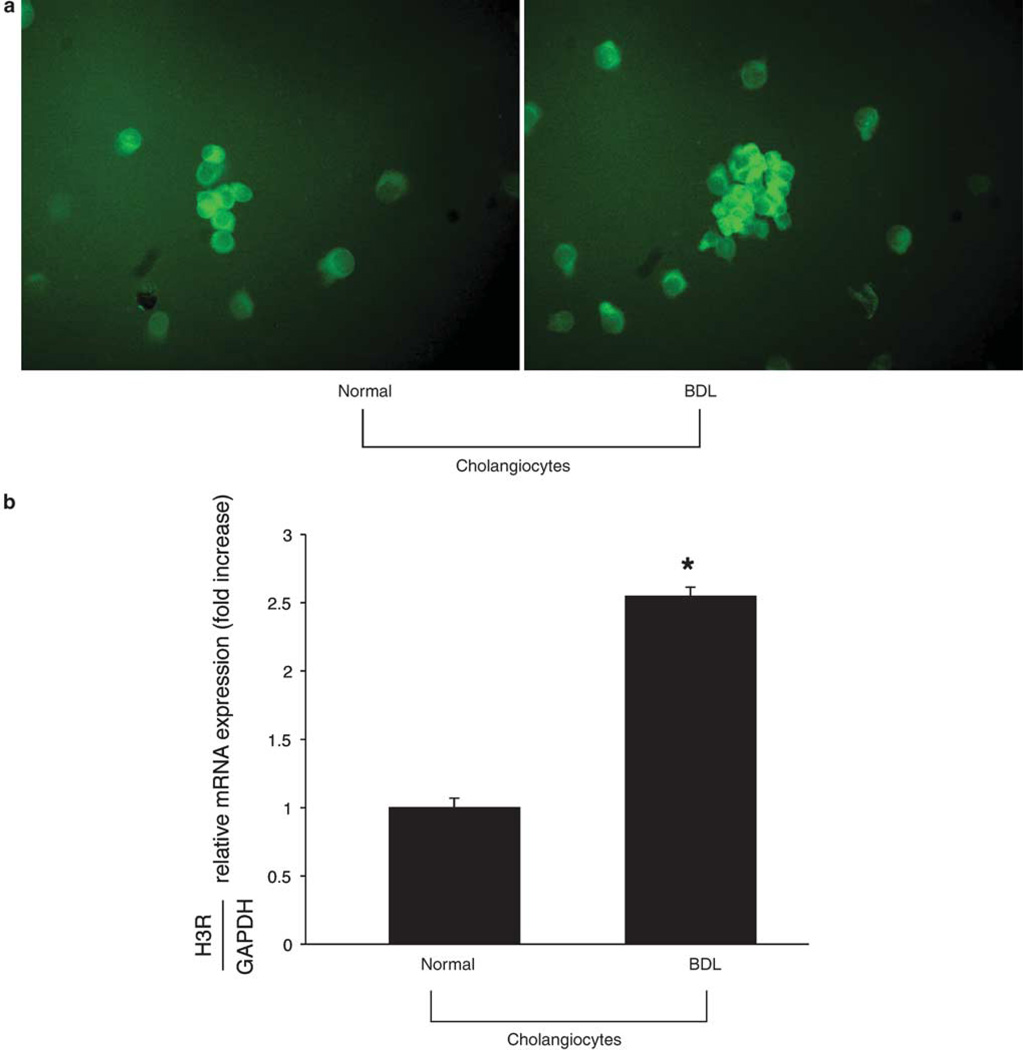
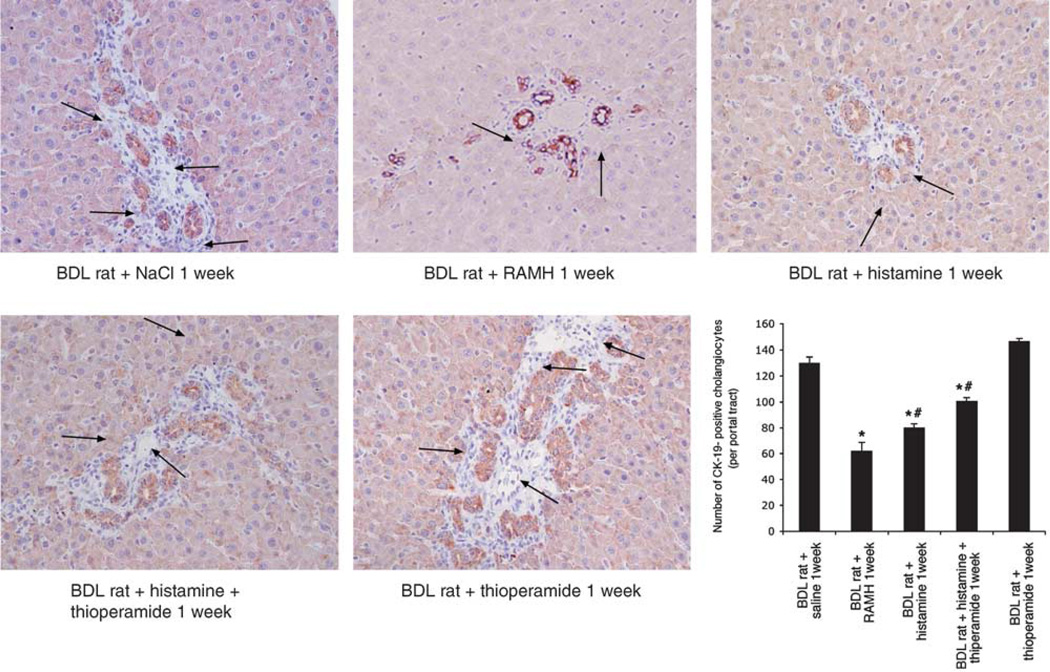
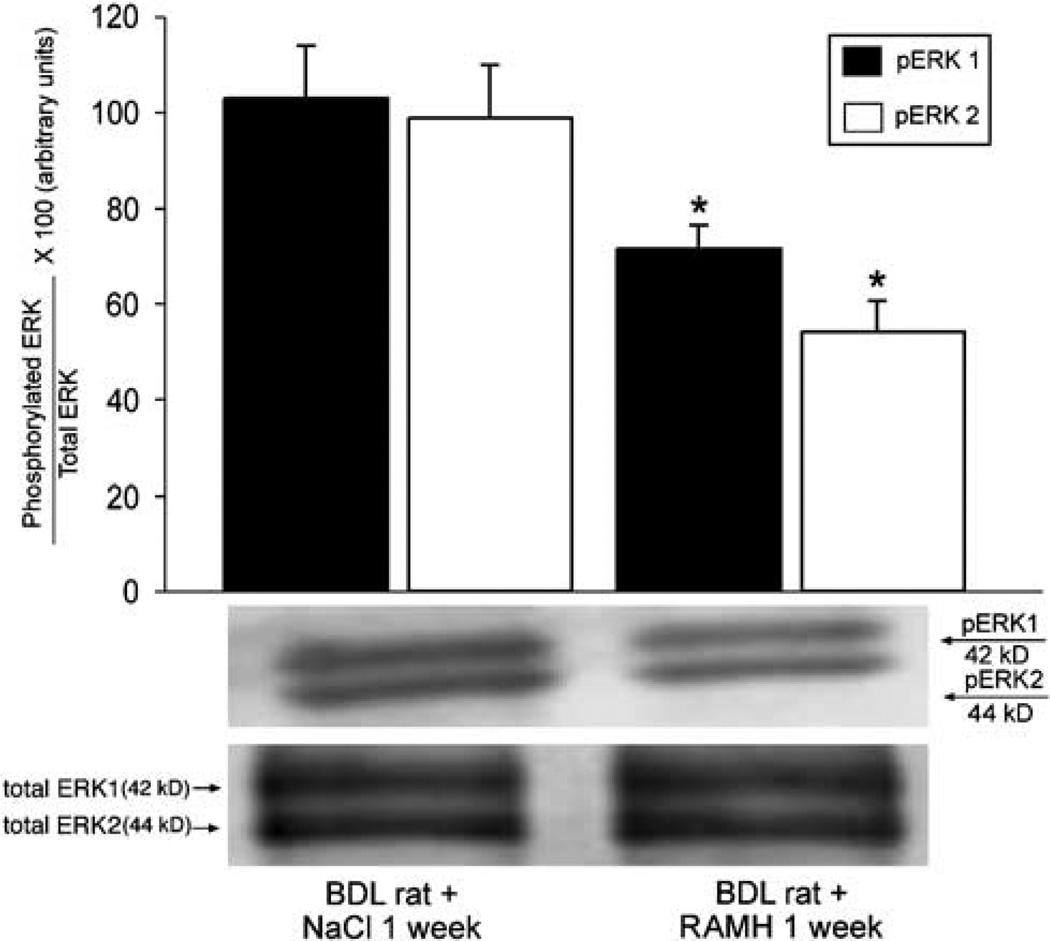
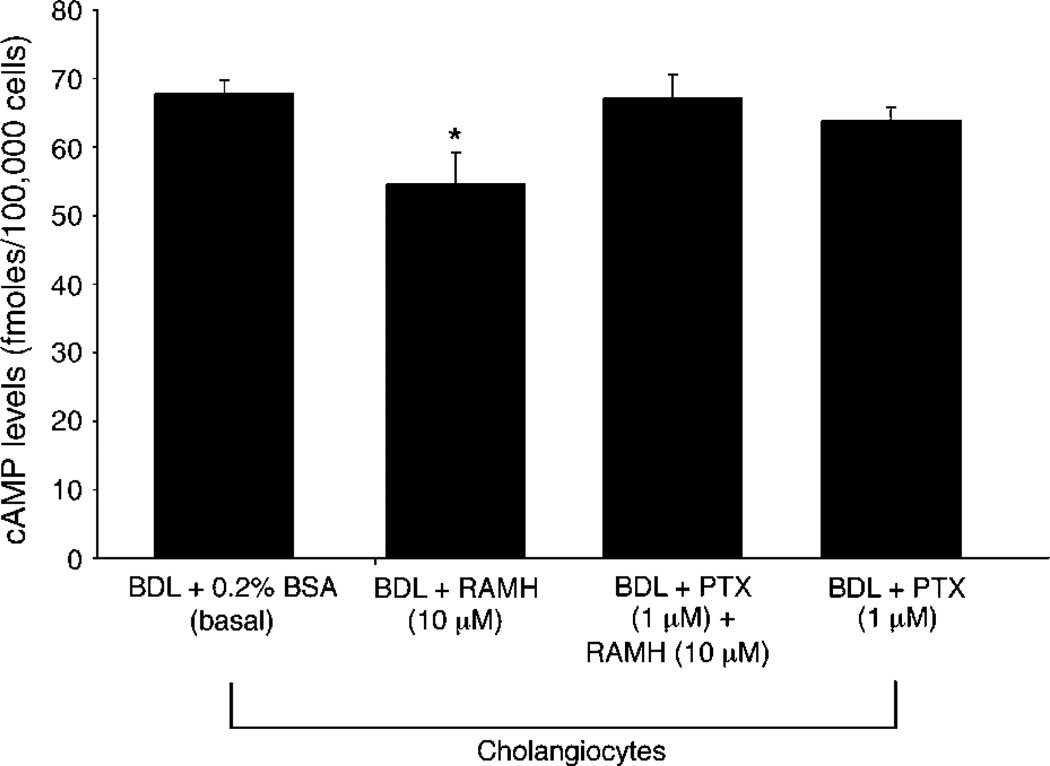
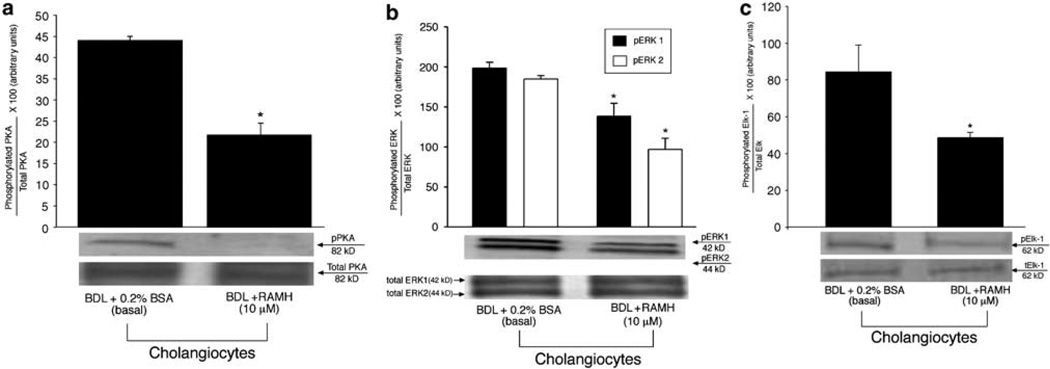
Histamine regulates many functions by binding to four histamine G-coupled receptor proteins (H1R, H2R, H3R and H4R). As H3R exerts their effects by coupling to Galpha(i/o) proteins reducing adenosine 3', 5'-monophosphate (cAMP) levels (a key player in the modulation of cholangiocyte hyperplasia/damage), we evaluated the role of H3R in the regulation of biliary growth. We posed the following questions: (1) Do cholangiocytes express H3R? (2) Does in vivo administration of (R)-(alpha)-(-)-methylhistamine dihydrobromide (RAMH) (H3R agonist), thioperamide maleate (H3R antagonist) or histamine, in the absence/presence of thioperamide maleate, to bile duct ligated (BDL) rats regulate cholangiocyte proliferation? and (3) Does RAMH inhibit cholangiocyte proliferation by downregulation of cAMP-dependent phosphorylation of protein kinase A (PKA)/extracellular signal-regulated kinase 1/2 (ERK1/2)/ets-like gene-1 (Elk-1)? The expression of H3R was evaluated in liver sections by immunohistochemistry and immunofluorescence, and by real-time PCR in cholangiocyte RNA from normal and BDL rats. BDL rats (immediately after BDL) were treated daily with RAMH, thioperamide maleate or histamine in the absence/presence of thioperamide maleate for 1 week. Following in vivo treatment of BDL rats with RAMH for 1 week, and in vitro stimulation of BDL cholangiocytes with RAMH, we evaluated cholangiocyte proliferation, cAMP levels and PKA, ERK1/2 and Elk-1 phosphorylation. Cholangiocytes from normal and BDL rats express H3R. The expression of H3R mRNA increased in BDL compared to normal cholangiocytes. Histamine decreased cholangiocyte growth of BDL rats to a lower extent than that observed in BDL RAMH-treated rats; histamine-induced inhibition of cholangiocyte growth was partly blocked by thioperamide maleate. In BDL rats treated with thioperamide maleate, cholangiocyte hyperplasia was slightly higher than that of BDL rats. In vitro, RAMH inhibited the proliferation of BDL cholangiocytes. RAMH inhibition of cholangiocyte growth was associated with decreased cAMP levels and PKA/ERK1/2/Elk-1 phosphorylation. Downregulation of cAMP-dependent PKA/ERK1/2/Elk-1 phosphorylation (by activation of H3R) is important in the inhibition of cholangiocyte growth in liver diseases.
Figures





References
-
- Alvaro D, Mancino MG, Glaser S, et al. Proliferating cholangiocytes: a neuroendocrine compartment in the diseased liver. Gastroenterology. 2006;132:415–431. - PubMed
-
- LeSage G, Glaser S, Alpini G. Regulation of cholangiocyte proliferation. Liver. 2001;21:73–80. - PubMed
-
- Alpini G, Prall RT, LaRusso NF. The pathobiology of biliary epithelia. In: Arias IM, Boyer JL, Chisari FV, Fausto N, Jakoby W, Schachter D, Shafritz DA, editors. The Liver, Biology and Pathobiology. 4th edn. Philadelphia, PA: Lippincott Williams & Wilkins; 2001. pp. 421–435.
-
- Francis H, Glaser S, Ueno Y, et al. cAMP stimulates the secretory and proliferative capacity of the rat intrahepatic biliary epithelium through changes in the PKA/Src/MEK/ERK1/2 pathway. J Hepatol. 2004;41:528–537. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
