

Review
doi: 10.1016/j.febslet.2007.01.094.
Epub 2007 Feb 20.
Regulating cytoskeleton-based vesicle motility
Affiliations
- PMID: 17335816
- PMCID: PMC1974873
- DOI: 10.1016/j.febslet.2007.01.094
Item in Clipboard
Review
Regulating cytoskeleton-based vesicle motility
FEBS Lett.
.
Abstract
During vesicular transport, the assembly of the coat complexes and the selection of cargo proteins must be coordinated with the subsequent translocation of vesicles from the donor to an acceptor compartment. Here, we review recent progress toward uncovering the molecular mechanisms that connect transport vesicles to the protein machinery responsible for cytoskeleton-mediated motility. An emerging theme is that vesicle cargo proteins, either directly or through binding interactions with coat proteins, are able to influence cytoskeletal dynamics and motor protein function. Hence, a vesicle's cargo composition may help direct its intracellular motility and targeting.
Figures
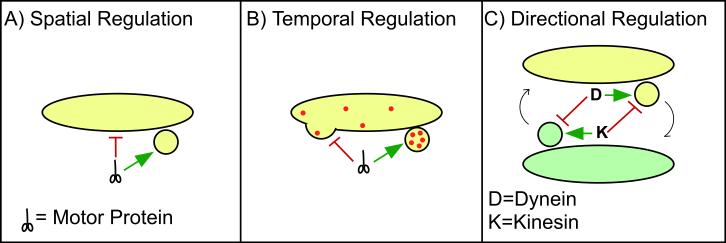
(A) Mechanisms for spatial regulation must exist to ensure that while a nascent vesicle binds to a motor and becomes motile, the donor organelle does not. (B) Temporal regulation should occur in order to coordinate motor recruitment with other steps in vesicle formation. Motor-based motility should be blocked until vesicle coat assembly, cargo packaging, and vesicle scission have been completed (C) In the cases where vesicles or cargo are recycled or undergo bidirectional transport, regulatory processes must ensure that the correct motor is functional. For example, during recycling between the ER and the Golgi apparatus coatomer-coated vesiculotubular clusters undergo dynein-mediated anterograde transport, whereas coatomer-coated COPI vesicles mediating retrograde transport from the Golgi apparatus utilize kinesin motors.
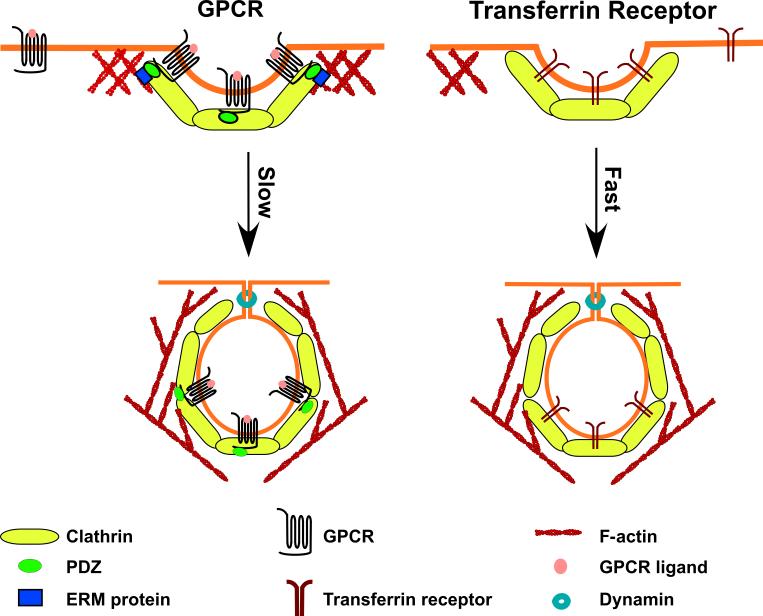
G-protein coupled receptors (GPCRs) are recruited to clathrin-coated pits after ligand binding (Left). For GPCRs that contain PDZ-ligand domains, an anchor between the forming vesicle and the actin cytoskeleton may be formed via PDZ-domain-containing proteins and the ERM-family of actin binding proteins. This creates clathrin-coated pits with delayed internalization and extended cell-surface residence times [22]. The transferrin receptor (Right) is also internalized through clathrin-coated pits. However, transferrin lacks a PDZ domain and does not form ERM-dependent interactions with cortical actin. Thus for transferring containing vesicles, the scission and internalization steps proceed rapidly.
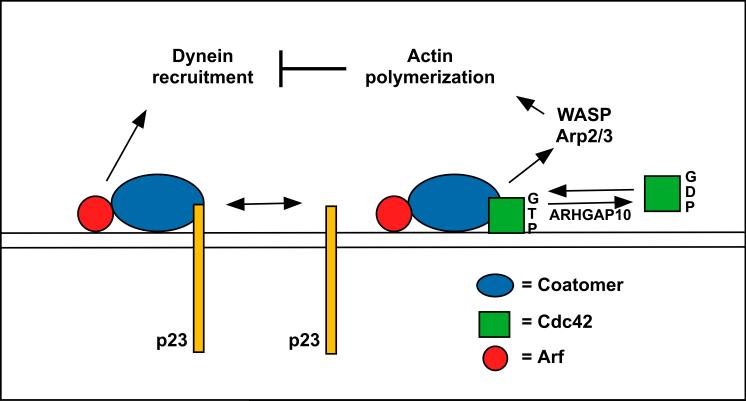
Actin polymerization on Golgi vesicles is stimulated by the ARF1-dependent recruitment of a complex between the COPI-coat protein, coatomer and the Rho-family GTP-binding protein, Cdc42 [35]. ARF1-dependent Cdc42 function at the Golgi apparatus is specifically regulated by the GTPase activating protein, ARHGAP10 [39]. Active Cdc42 also inhibits dynein recruitment in an actin-dependent manner [36]. Coatomer cannot be simultaneously bound to Cdc42 and the p23 putative cargo receptor. Thus, the presence of p23 acts to block actin polymerization and stimulate dynein recruitment. This signaling may ensure that vesicle motility does not commence until the completion of vesicle assembly and cargo packaging.
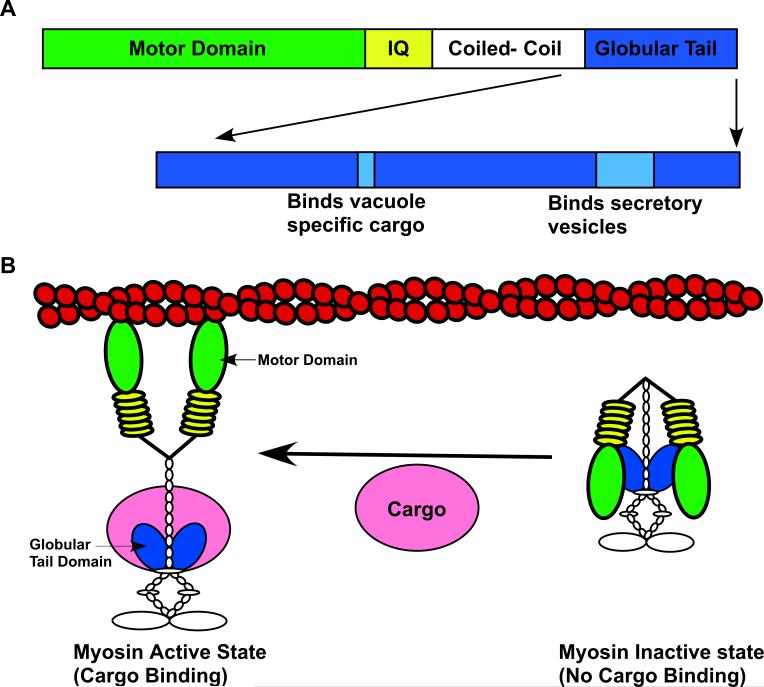
(A) The heavy chain of myosin 5 contains an N-terminal head domain and a C-terminal globular tail domain. The head domain contains the binding sites for actin and ATP. The globular tail domain contains the cargo-binding sites. Structural analysis of the yeast myosin 5 protein, Myo2p, reveals that the globular tail contains two distinct cargo-binding modules one for transport vesicles and a second for vacuoles [45,46]. (B) The globular tail of myosin 5a can bind and inhibit the ATPase activity of the N-terminal head domain. It is proposed that cargo binding to the globular tail domain of myosin 5 causes a conformation change leading to the activation of the motor [50,51].
Similar articles
-
Regulating the actin cytoskeleton during vesicular transport.Curr Opin Cell Biol. 2002 Aug;14(4):428-33. doi: 10.1016/s0955-0674(02)00349-6. Curr Opin Cell Biol. 2002. PMID: 12383793 Review.
-
Cargo takes control of endocytosis.Cell. 2006 Oct 6;127(1):35-7. doi: 10.1016/j.cell.2006.09.012. Cell. 2006. PMID: 17018271
-
Motoring around the Golgi.Nat Cell Biol. 2002 Oct;4(10):E236-42. doi: 10.1038/ncb1002-e236. Nat Cell Biol. 2002. PMID: 12360306 Review.
-
Coiled coils and SAH domains in cytoskeletal molecular motors.Biochem Soc Trans. 2011 Oct;39(5):1142-8. doi: 10.1042/BST0391142. Biochem Soc Trans. 2011. PMID: 21936779 Review.
-
Unconventional myosins: anchors in the membrane traffic relay.Traffic. 2000 Jan;1(1):11-8. doi: 10.1034/j.1600-0854.2000.010103.x. Traffic. 2000. PMID: 11208054 Review.
Cited by
-
Spatial modeling of vesicle transport and the cytoskeleton: the challenge of hitting the right road.PLoS One. 2012;7(1):e29645. doi: 10.1371/journal.pone.0029645. Epub 2012 Jan 12. PLoS One. 2012. PMID: 22253752 Free PMC article.
-
Galectin-9 interacts with Vamp-3 to regulate cytokine secretion in dendritic cells.Cell Mol Life Sci. 2023 Sep 27;80(10):306. doi: 10.1007/s00018-023-04954-x. Cell Mol Life Sci. 2023. PMID: 37755527 Free PMC article.
-
Identification of Transferrin Receptor 1 (TfR1) Overexpressed in Lung Cancer Cells, and Internalization of Magnetic Au-CoFe2O4 Core-Shell Nanoparticles Functionalized with Its Ligand in a Cellular Model of Small Cell Lung Cancer (SCLC).Pharmaceutics. 2022 Aug 17;14(8):1715. doi: 10.3390/pharmaceutics14081715. Pharmaceutics. 2022. PMID: 36015341 Free PMC article.
-
Actin acting at the Golgi.Histochem Cell Biol. 2013 Sep;140(3):347-60. doi: 10.1007/s00418-013-1115-8. Epub 2013 Jun 27. Histochem Cell Biol. 2013. PMID: 23807268 Review.
-
The role of actin remodeling in the trafficking of intracellular vesicles, transporters, and channels: focusing on aquaporin-2.Pflugers Arch. 2008 Jul;456(4):737-45. doi: 10.1007/s00424-007-0404-2. Epub 2007 Dec 8. Pflugers Arch. 2008. PMID: 18066585 Review.
References
-
- Caviston JP, Holzbaur EL. Microtubule motors at the intersection of trafficking and transport. Trends Cell Biol. 2006;16:530–7. - PubMed
-
- Soldati T, Schliwa M. Powering membrane traffic in endocytosis and recycling. Nat Rev Mol Cell Biol. 2006;7:897–908. - PubMed
-
- Egea G, Lazaro-Dieguez F, Vilella M. Actin dynamics at the Golgi complex in mammalian cells. Curr Opin Cell Biol. 2006;18:168–178. - PubMed
-
- Orci L, Stamnes M, Ravazzola M, Amherdt M, Perrelet A, Sollner TH, Rothman JE. Bidirectional transport by distinct populations of COPI-coated vesicles. Cell. 1997;90:335–49. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
