

Spinal CGRP1 receptors contribute to supraspinally organized pain behavior and pain-related sensitization of amygdala neurons
- PMID: 17335972
- PMCID: PMC2066202
- DOI: 10.1016/j.pain.2007.01.024
Spinal CGRP1 receptors contribute to supraspinally organized pain behavior and pain-related sensitization of amygdala neurons
Abstract
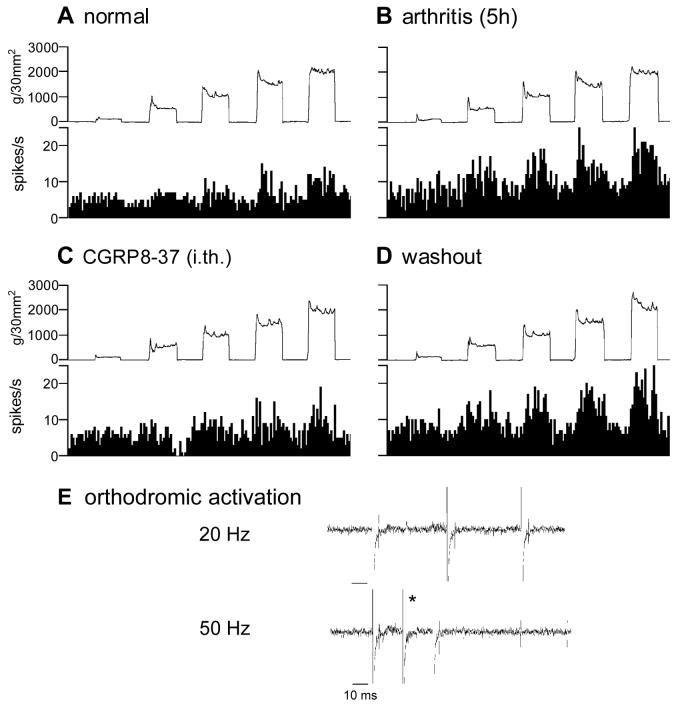
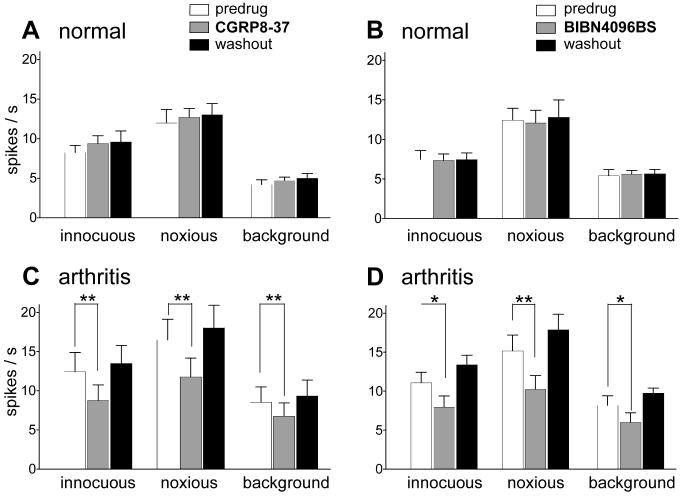
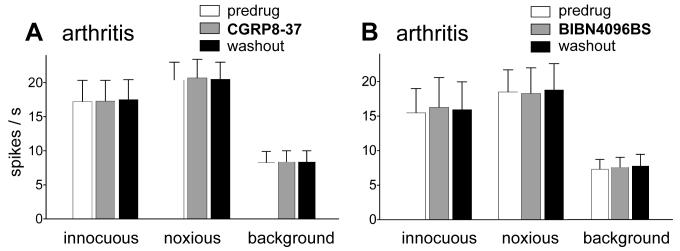
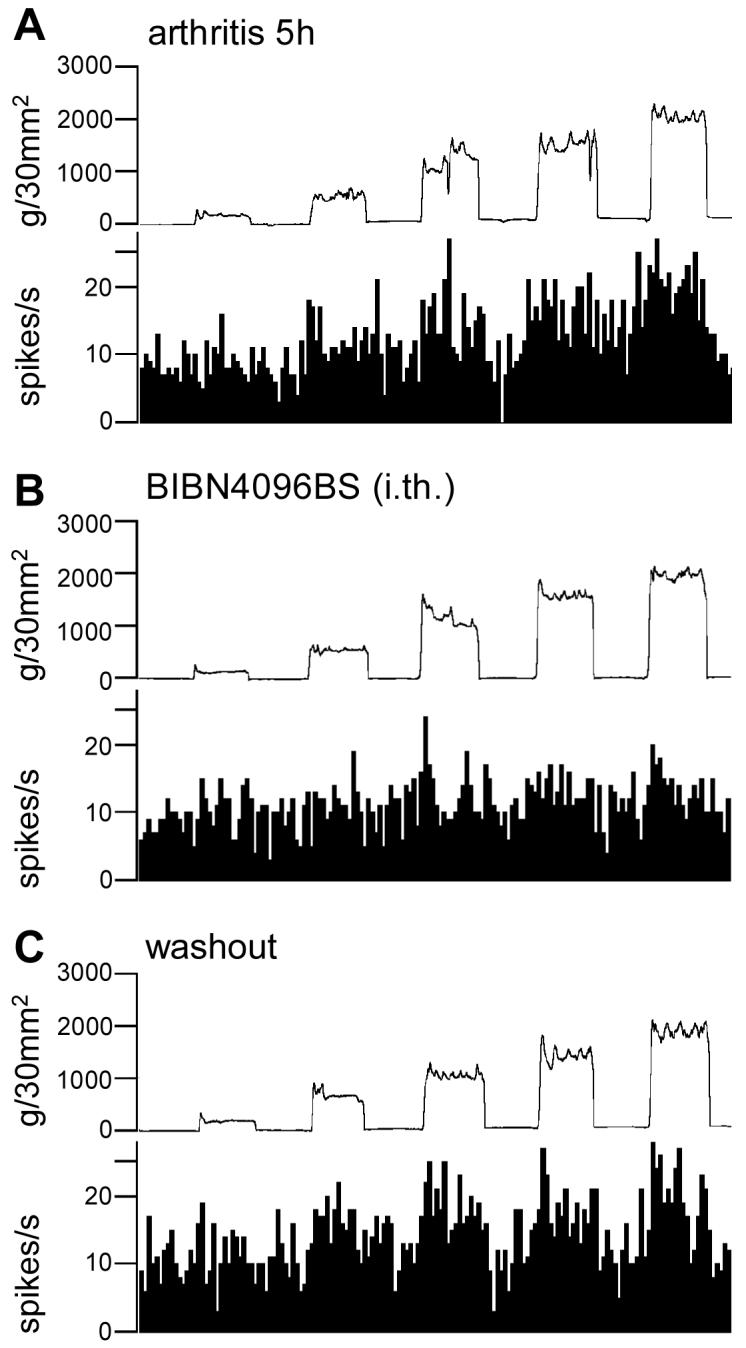
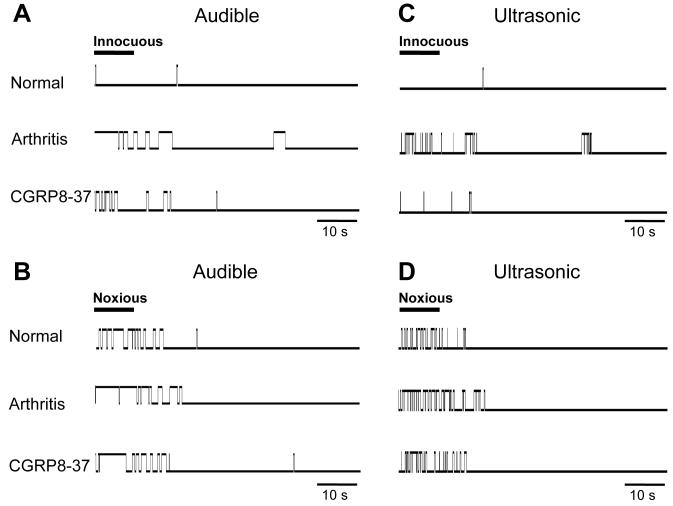
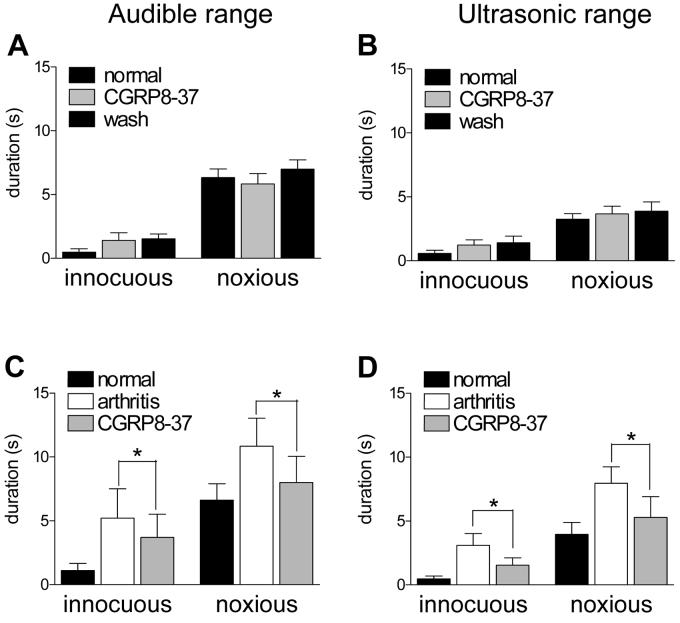
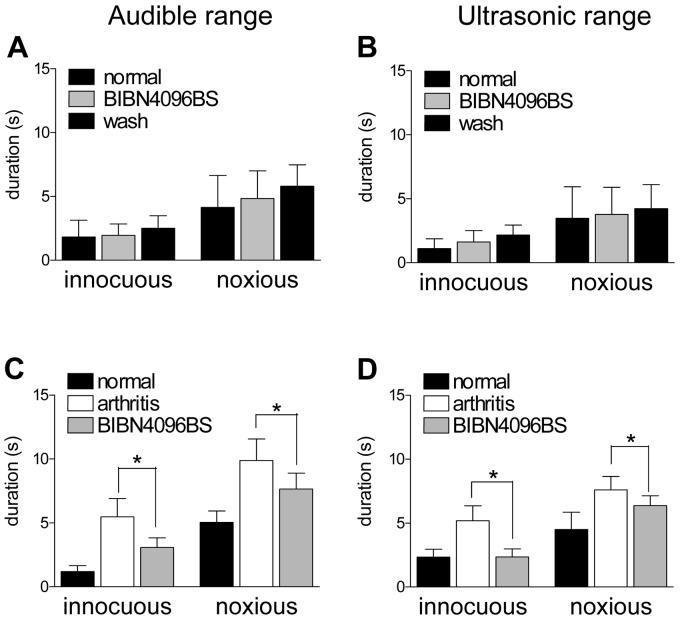
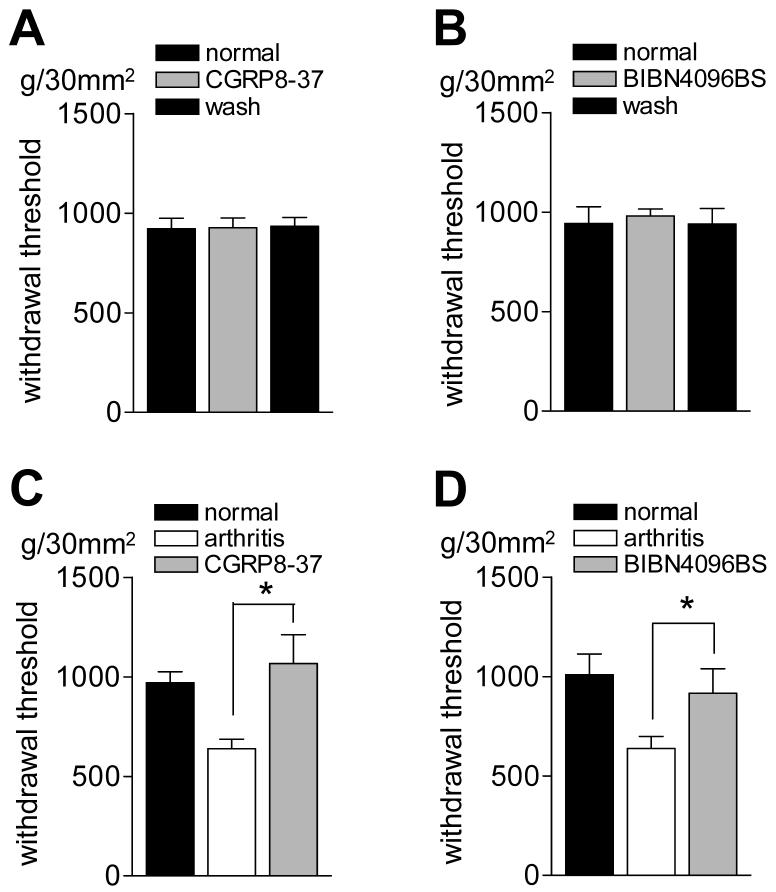
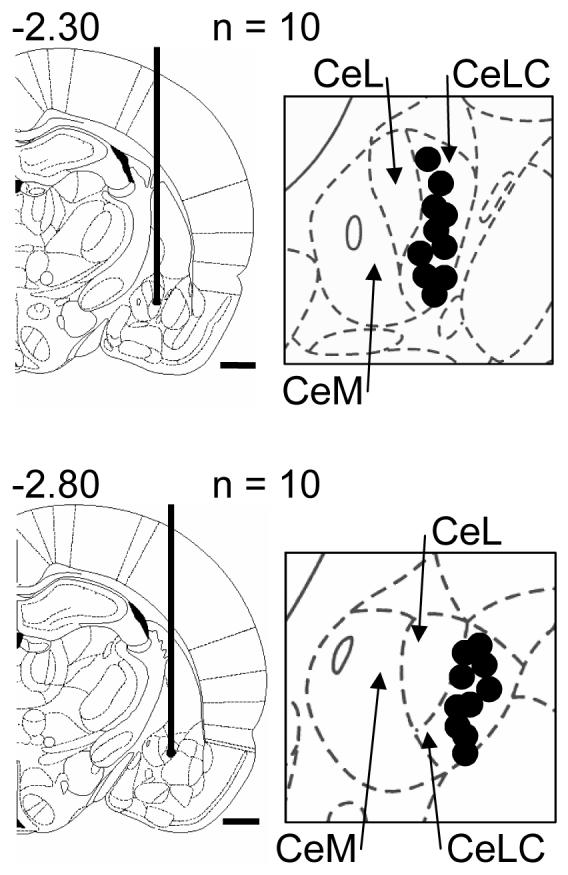
CGRP receptor activation has been implicated in peripheral and central sensitization. The role of spinal CGRP receptors in supraspinal pain processing and higher integrated pain behavior is not known. Here we studied the effect of spinal inhibition of CGRP1 receptors on supraspinally organized vocalizations and activity of amygdala neurons. Our previous studies showed that pain-related audible and ultrasonic vocalizations are modulated by the central nucleus of the amygdala (CeA). Vocalizations in the audible and ultrasonic range and hindlimb withdrawal thresholds were measured in awake adult rats before and 5-6h after induction of arthritis by intra-articular injections of kaolin and carrageenan into one knee. Extracellular single-unit recordings were made from neurons in the latero-capsular division of the CeA (CeLC) in anesthetized rats before and after arthritis induction. CGRP1 receptor antagonists were applied to the lumbar spinal cord intrathecally (5 microl/min) 6h postinduction of arthritis. Spinal administration of peptide (CGRP8-37, 1 microM) and non-peptide (BIBN4096BS, 1 microM) CGRP1 receptor antagonists significantly inhibited the increased responses of CeLC neurons to mechanical stimulation of the arthritic knee but had no effect under normal conditions. In arthritic rats, the antagonists also inhibited the audible and ultrasonic components of vocalizations evoked by noxious stimuli and increased the threshold of hindlimb withdrawal reflexes. The antagonists had no effect on vocalizations and spinal reflexes in normal rats. These data suggest that spinal CGRP1 receptors are not only important for spinal pain mechanisms but also contribute significantly to the transmission of nociceptive information to the amygdala and to higher integrated behavior.
Figures
References
-
- Bernard J-F, Bester H, Besson JM. Involvement of the spino-parabrachio -amygdaloid and -hypothalamic pathways in the autonomic and affective emotional aspects of pain. Prog Brain Res. 1996;107:243–255. - PubMed
-
- Borszcz GS. Increases in vocalization and motor reflex thresholds are influenced by the site of morphine microinjection: comparisons following administration into the periaqueductal gray, ventral medulla, and spinal subarachnoid space. Behav Neurosci. 1995;109:502–522. - PubMed
-
- Borszcz GS, Johnson CP, Williams DH. Increases in vocalization and motor reflex thresholds generated by the intrathecal administration of serotonin or norepinephrine. Behav Neurosci. 1996;110:809–822. - PubMed
-
- Borszcz GS, Leaton RN. The effect of amygdala lesions on conditional and unconditional vocalizations in rats. Neurobiol Learn and Mem. 2003;79:212–225. - PubMed
-
- Borta A, Wohr M, Schwarting RKW. Rat ultrasonic vocalization in aversively motivated situations and the role of individual differences in anxiety-related behavior. Behavioural Brain Research. 2006;166:271–280. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
