

Actin is involved in auxin-dependent patterning
- PMID: 17337532
- PMCID: PMC1851825
- DOI: 10.1104/pp.106.094052
Actin is involved in auxin-dependent patterning
Abstract
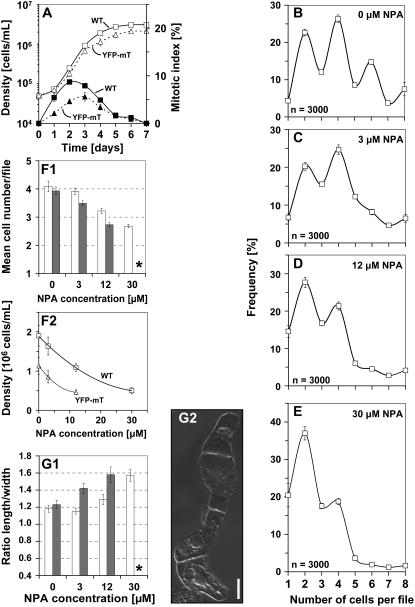
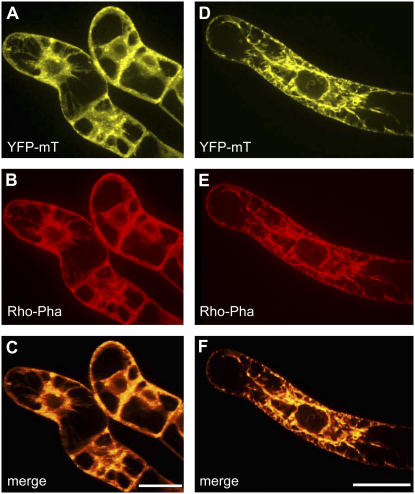
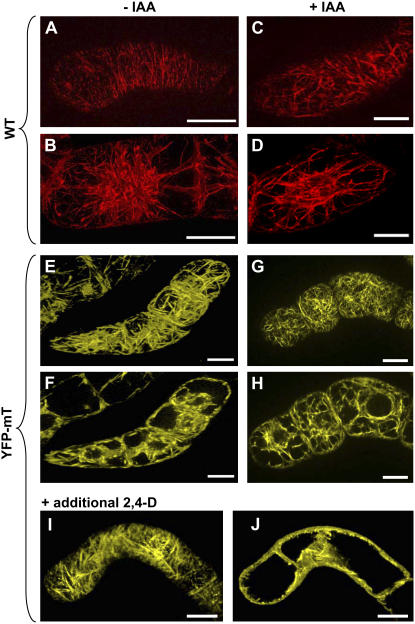
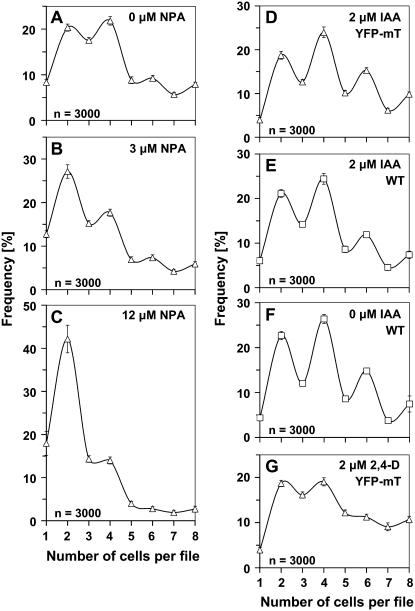
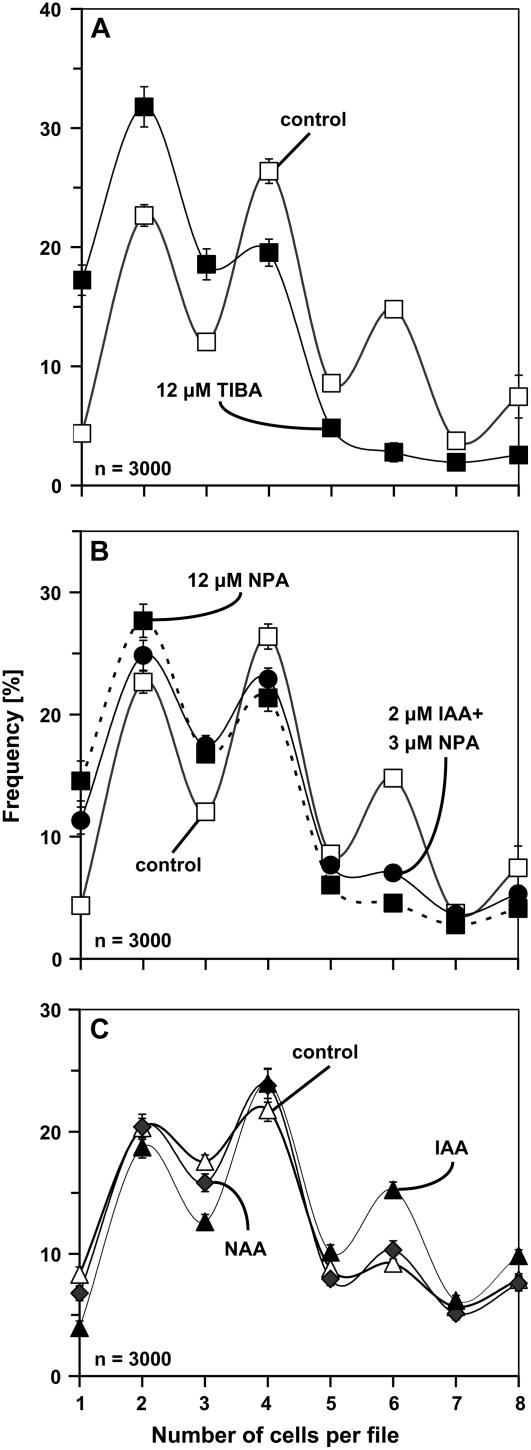
Polar transport of auxin has been identified as a central element of pattern formation. The polarity of auxin transport is linked to the cycling of pin-formed proteins, a process that is related to actomyosin-dependent vesicle traffic. To get insight into the role of actin for auxin transport, we used patterned cell division to monitor the polarity of auxin fluxes. We show that cell division in the tobacco (Nicotiana tabacum L. cv Bright-Yellow 2) cell line is partially synchronized and that this synchrony can be perturbed by inhibition of auxin transport by 1-N-naphthylphthalamic acid. To address the role of actin in this synchrony, we induced a bundled configuration of actin by overexpressing mouse talin. The bundling of actin impairs the synchrony of cell division and increases the sensitivity to 1-N-naphthylphthalamic acid. Addition of the polarly transported auxins indole-3-acetic acid and 1-naphthyl acetic acid (but not 2,4-dichlorophenoxyacetic acid) restored both the normal organization of actin and the synchrony of cell division. This study suggests that auxin controls its own transport by changing the state of actin filaments.
Figures
Similar articles
-
Nicotiana tabacum actin-depolymerizing factor 2 is involved in actin-driven, auxin-dependent patterning.J Plant Physiol. 2013 Aug 15;170(12):1057-66. doi: 10.1016/j.jplph.2013.03.002. Epub 2013 Mar 30. J Plant Physiol. 2013. PMID: 23545293
-
Auxin stimulates its own transport by shaping actin filaments.Plant Physiol. 2009 Sep;151(1):155-67. doi: 10.1104/pp.109.140111. Epub 2009 Jul 24. Plant Physiol. 2009. PMID: 19633235 Free PMC article.
-
Light can rescue auxin-dependent synchrony of cell division in a tobacco cell line.J Exp Bot. 2010;61(2):503-10. doi: 10.1093/jxb/erp319. Epub 2009 Oct 30. J Exp Bot. 2010. PMID: 19884227 Free PMC article.
-
Probing the actin-auxin oscillator.Plant Signal Behav. 2010 Feb;5(2):94-8. doi: 10.4161/psb.5.2.10337. Epub 2010 Feb 15. Plant Signal Behav. 2010. PMID: 20023411 Free PMC article. Review.
-
Keeping it all together: auxin-actin crosstalk in plant development.J Exp Bot. 2015 Aug;66(16):4983-98. doi: 10.1093/jxb/erv308. Epub 2015 Jun 17. J Exp Bot. 2015. PMID: 26085676 Review.
Cited by
-
Switching cell fate by the actin-auxin oscillator in Taxus: cellular aspects of plant cell fermentation.Plant Cell Rep. 2022 Dec;41(12):2363-2378. doi: 10.1007/s00299-022-02928-0. Epub 2022 Oct 10. Plant Cell Rep. 2022. PMID: 36214871 Free PMC article.
-
Defence signalling triggered by Flg22 and Harpin is integrated into a different stilbene output in Vitis cells.PLoS One. 2012;7(7):e40446. doi: 10.1371/journal.pone.0040446. Epub 2012 Jul 6. PLoS One. 2012. PMID: 22792328 Free PMC article.
-
Myosins XI-K, XI-1, and XI-2 are required for development of pavement cells, trichomes, and stigmatic papillae in Arabidopsis.BMC Plant Biol. 2012 Jun 6;12:81. doi: 10.1186/1471-2229-12-81. BMC Plant Biol. 2012. PMID: 22672737 Free PMC article.
-
Tobacco Arp3 is localized to actin-nucleating sites in vivo.J Exp Bot. 2009;60(2):603-14. doi: 10.1093/jxb/ern307. Epub 2009 Jan 6. J Exp Bot. 2009. PMID: 19129161 Free PMC article.
-
Auxin influx inhibitors 1-NOA, 2-NOA, and CHPAA interfere with membrane dynamics in tobacco cells.J Exp Bot. 2010 Aug;61(13):3589-98. doi: 10.1093/jxb/erq172. Epub 2010 Jul 1. J Exp Bot. 2010. PMID: 20595238 Free PMC article.
References
-
- Berleth T, Sachs T (2001) Plant morphogenesis: long-distance coordination and local patterning. Curr Opin Plant Biol 4 57–62 - PubMed
-
- Blakeslee JJ, Peer WA, Murphy AS (2005) Auxin transport. Curr Opin Plant Biol 8 494–500 - PubMed
-
- Chen R, Masson PH (2006) Auxin transport and recycling of PIN proteins in Plants. In J Šamaj, F Baluška, D Menzel, eds, Plant Endocytosis. Springer, Berlin, pp 139–157
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
